The Dread Olecranon of Kentrosaurus
July 4, 2025
(This was buried in Part 5 of my 2011 review of the Sideshow Apatosaurus maquette, but it’s long deserved to be a post of its own, and now it is. I’m not adding anything new here, just extracting and reposting the relevant bits, for reasons that will become clear in a future post. — MJW)
The Dread Olecranon of Kentrosaurus is something Heinrich Mallison pointed out in the second of his excellent Plateosaurus papers (Mallison 2010: fig. 3).

Heinrich’s thoughts on articular cartilage in dinosaurs are well worth reading, so once again I’m going to quote extensively (Mallison 2010: p. 439):
Cartilaginous tissues are rarely preserved on fossils, so the thickness of cartilage caps in dinosaurs is unclear. Often, it is claimed that even large dinosaurs had only thin layers of articular cartilage, as seen in extant large mammals, because layers proportional to extant birds would have been too thick to be effectively supplied with nutrients from the synovial fluid. This argument is fallacious, because it assumes that a thick cartilage cap on a dinosaur long bone would have the same internal composition as the thin cap on a mammalian long bone. Mammals have a thin layer of hyaline cartilage only, but in birds the structure is more complex, with the hyaline cartilage underlain by thicker fibrous cartilage pervaded by numerous blood vessels (Graf et al. 1993: 114, fig. 2), so that nutrient transport is effected through blood vessels, not diffusion. This tissue can be scaled up to a thickness of several centimeters without problems.
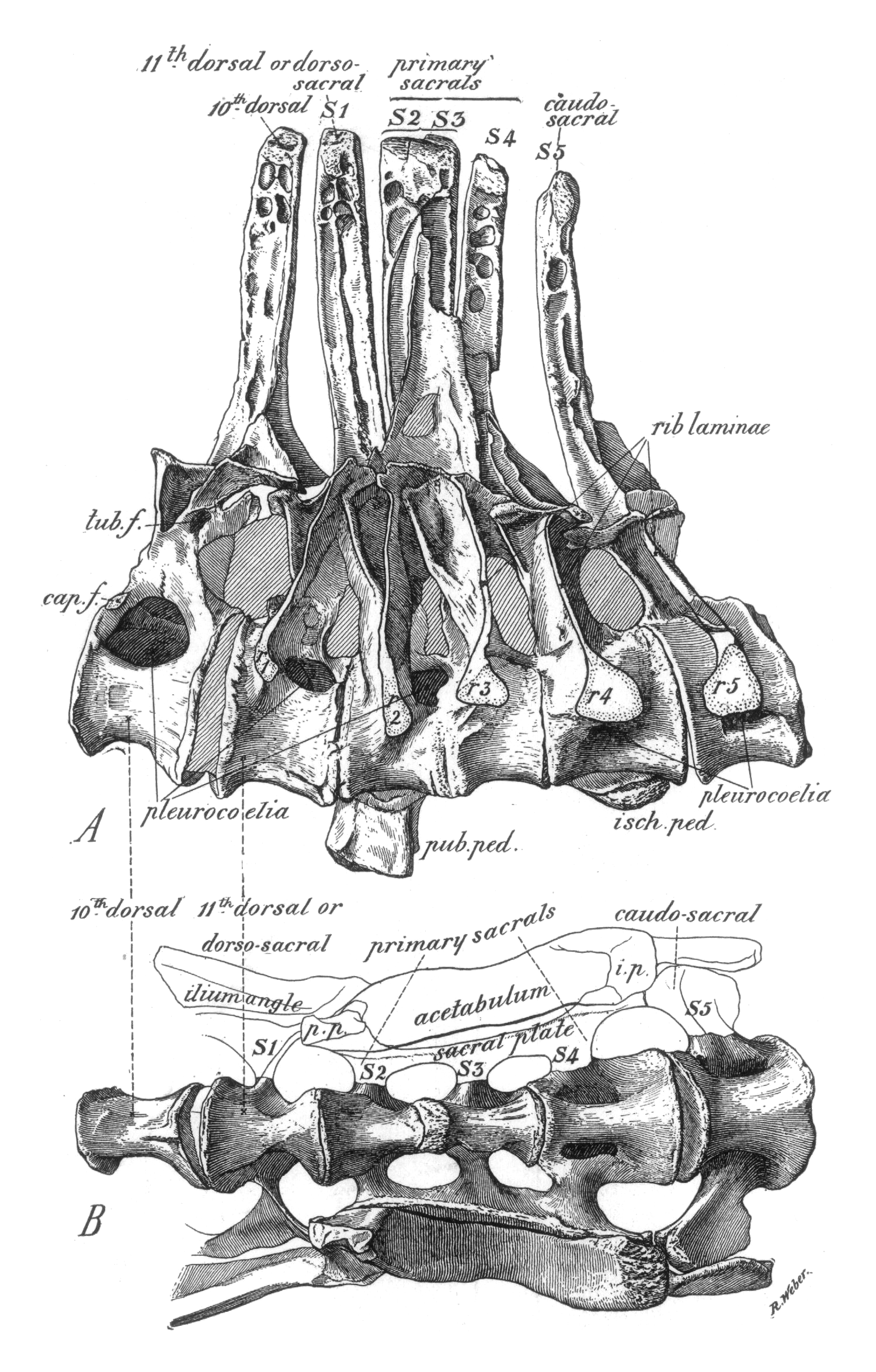
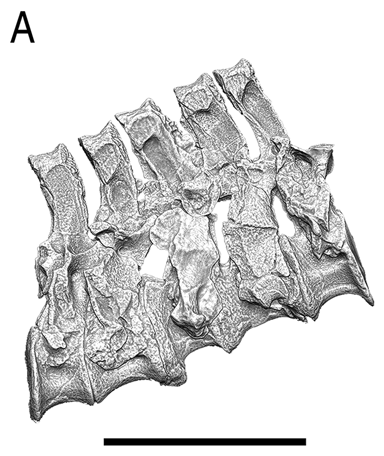
An impressive example for the size of cartilaginous structures in dinosaurs is the olecranon process in the stegosaur Kentrosaurus aethiopicus Hennig, 1915. In the original description a left ulna (MB.R.4800.33, field number St 461) is figured (Hennig 1915: fig. 5) that shows a large proximal process. However, other ulnae of the same species lack this process, and are thus far less distinct from other dinosaurian ulnae (Fig. 3B, C). The process on MB.R.4800.33 and other parts of its surface have a surface texture that can also be found on other bones of the same individual, and may indicate some form of hyperostosis or another condition that leads to ossification of cartilaginous tissues. Fig. 3B–D compares MB.R.4800.33 and two other ulnae of K. aethiopicus from the IFGT skeletal mount. It is immediately obvious that the normally not fossilized cartilaginous process has a significant influence on the ability to hyperextend the elbow, because it forms a stop to extension. Similarly large cartilaginous structures may have been present on a plethora of bones in any number of dinosaur taxa, so that range of motion analyses like the one presented here are at best cautious approximations.
One of the crucial points to take away from all of this is that thick cartilage caps did not only expand or only limit the ranges of motions of different joints. The mistake is to think that soft tissues always do one or the other. The big olecranon in Kentrosaurus probably limited the ROM of the elbow, by banging into the humerus in extension. In contrast, thick articular cartilage at the wrists [of sauropods] probably expanded the ROM and may have allowed the strong wrist flexion that some artists have restored for sauropods. I’m not arguing that it must have done so, just that I don’t think we can rule out the possibility that it may have.
– – – – – – – – – – – – – – – – – – – –
To see that chunk in context and read more about cartilage in dinosaurs, see the original post — here’s that link again. Throwing in the references to Bonnan et al. (2010) and Holliday et al. (2010) because they’re still relevant, foundational studies.
References
- Bonnan, M.F., Sandrik, J.L., Nishiwaki, T., Wilhite, D.R., Elsey, R.M., and Vittore, C. 2010. Calcified cartilage shape in archosaur long bones reflects overlying joint shape in stress-bearing elements: Implications for nonavian dinosaur locomotion. The Anatomical Record 293: 2044-2055.
- Holliday, C.M., R.C. Ridgely, J.C. Sedlmayr and L.M. Witmer. 2010. Cartilaginous epiphyses in extant archosaurs and their implications for reconstructing limb function in dinosaurs. PLoS ONE 5(9): e13120. doi:10.1371/journal.pone.0013120
- Mallison, H. 2010. The digital Plateosaurus II: An assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount. Acta Palaeontologica Polonica 55 (3): 433–458.
Five questions from Tom Redd
October 24, 2024
Three years ago, Tom Redd made a very generous commitment to the SV-POW! Patreon, and he remains our most generous donor in total. When I wrote to thank him his reply included “I have thousands of questions about apatosaurus that I would like to ask you some day.”
It seemed only fair to invite him to ask some of those questions, so we asked him to give us five and said we’d try to answer them. When the questions came through, some of them were hard — not really in our area of expertise. But I promised we’d take a crack, and that we’d invite commenters to chip in where we get something wrong or leave something out.
Then a bit more than three years slipped past. Now, finally: here we go!
Question #1:
Do you think sauropods could have evolved long necks as a defensive strategy? (Better to see your enemies from a long way off). The sigmoid curve and the neutral position is another issue!
I recently read a couple of research papers where the researcher used “radiological imaging” to determine the neutral position of the cervical vertebrae. The result was a gently downward curving neck beginning at the pectoral girdle, with the skull only 2 meters off the ground. (I agree with Matt that this would be an ambush predator’s delight!)
Also, could compressive or tensional forces of cartilage and ligaments affect the neutral position of the cervical vertebra? (I believe Matt also alluded to this condition.)
This is the longest question, but maybe the one I’m best positioned to comment on.
First of all, the perennial question of what sauropods’ long necks were for. There are a few candidate explanations out there. The obvious one is that they enabled high browsing, and on the whole we feel that’s the strongest single explanation. Another candidate factor is sexual selection — that sauropods were particularly attracted to long-necked individuals of the opposite sex — but we don’t feel that is a strong explanation for reasons explained in our 2011 paper (Taylor et al. 2011). Predator avoidance would certainly have been aided by the long visual distances allowed by elevated heads, and we are confident that at least some sauropods used their necks in combat — primarily in intraspecific combat (Taylor 2015) but no doubt also against predators when the occasion arose. And we may well have missed other good uses for long necks.
In reality of course all these factors likely played a role: structures do not always, or even often, evolve for a single reason. When people who know much more about ceratopsians tell me “The horns of Triceratops were for intraspecific display and combat”, I don’t doubt them. But I also don’t doubt that, whatever the primary purpose of the horns, a Triceratops confronted by a Tyrannosaurus would do its damnedest to stick its horns into it. In the same way, while high feeding seems like the strongest driver of sauropod neck elongation, the other factors will surely have played in, too.
I’m not sure what radiological imaging papers you have come across, but the one I know about is Berman and Rothschild (2005) in the Thunder Lizards edited volume. This paper rather questionably partitions all sauropod cervicals into two bins, “robust” and “gracile”, and concludes, based on functional stress analysis, that “the robust-type centrum supported a neck held in a vertical, or near-vertical, pose, whereas the gracile-type centrum supported a neck held in a horizontal, or near-horizontal, pose”. If this is right, and their categorization holds, then Camarasaurus and an unidentified titanosaur had vertical necks; and Diplodocus, Apatosaurus, Haplocanthosaurus, Barosaurus and Brachiosaurus all had horizontal necks. We find every part of this unconvincing. At some point we should explain why in detail; but it is not this day.
Finally, yes, compressive and tensile forces in cartilage and ligaments definitely did affect neutral posture. My 2014 paper (Taylor 2014) shows this rather dramatically.

What we’re seeing here is what the neutral posture would be if cartilage is added to a neck that is otherwise articulated in horizonzal pose. The importance of intervertebral cartilage has often been overlooked, but can make a dramatic difference to neck posture.
Question #2
I recently read a report that indicated Apatosaurus survived at the species level for a period of approximately 8 million years ! So is this a success, average, or a short run?
I’m not sure where you read that, but the problem here is that no-one really knows what Apatosaurus means. We have the type species Apatosaurus ajax, sure, and the referred species Apatosaurus louisae, and the genus Brontosaurus based on the species Brontosaurus excelsus which is sometimes but not always synonymised with the genus Apatosaurus yielding the combination Apatosaurus excelsus, and don’t even get me started on Apatosaurus parvus, Apatosaurus laticollis, Atlantosaurus and whatever the heck AMNH 460 is.
So if we say that Apatosaurus survived for 8 million years, what exactly are we saying? That Apatosaurus ajax is known from sediments that differ in age by 8 million years? That would be interesting if true, but it’s very hard to establish because the referral of any given individual to a particular sauropod species tends to be very uncertain — largely because most specimen are so fragmentary and distorted. And if all we mean is that 8 million years separate the oldest and youngest specimens that have been referred to the genus Apatosaurus — well, that statement is all but meaningless, given the huge uncertainty about what is and is not part of that genus, if indeed genera even really mean anything.
Putting it all together, I’m not confident that there is any reason to think that Apatosaurus was particularly longer lived than other sauropods. Probably Camarasaurus outlasted it if you include all the taxa that have been referred to Camarasaurus. But then I’m far from convinced that that’s the right thing to do, too.
Question #3
Why so many heavy predators during the age of apatosaurs? Predator to prey ratios were in the 6 to 8% range as compared to modern ratios of 2 to 3%!
I’m not sure I can say much about this without knowing the source of the figures, but I assume that what’s being counted here is the number of individuals represented in the fossil record, and the ratio of a predator species to prey species. The problem is that there is a huge amount of vagueness in these numbers, but it’s not obvious that the apatosaur-age figures are comparable to the modern ones.
Consider first the modern ratio. Which animals are counted in each category? In the Serengeti, lions prey on zebras. So far, so simple, but there are also dwarf moongooses, which are predators — but they don’t hunt zebras. So do we count them in the numbers? If so, do we count their prey animals, too? Including invertebrates? And if not, then where do we draw the line between predators that we consider do and do not hunt the prey animals that we’ve decided we’re interested in?
Then there’s the matter of which animals get counted. If you do your Serengeti counts on dead animals, you might find disproportionately many predators because prey animals tend to be consumed. Or you might find disproportionately many prey animals because they tend to die in areas where the corpses are more easily found and counted. You might be able to do better by counting live animals, but then you might easily undercount secretive predators, or perhaps overcount predators because they stand fearlessly around to be counted.
Now consider trying to measure the predator/prey ratio in the Morrison formation. You have all the problems I already mentioned, plus a bunch of others. If you only count complete-ish articulated skeletons then your sample size is too small to be meaningful. If you count isolated elements, you’re at risk of registering multiple instances of the same individual. Counting individuals represented in bonebeds is difficult because of these problems. Assigning an element to a taxon is error-prone (though should generally be OK at the high level of sauropod vs. theropod — or are you?). Bones of different taxa may survive taphonomy better or worse than others. Life history differences will mean that the fossils of long-lived taxa under-represent their live populations. And so on, and on, and on.
Putting it all together, I would tend to be very sceptical that a difference in ratios of 6–8% to 2–3% is necessarily telling us anything.
With all that said, it’s perfectly possible that the average predator:prey body-size ratio was closer in the Morrison than in modern ecosystems. But we’d do better trying to measure that directly from body-fossils than to infer it from population densities.
Question #4
Are all apatosaur tracks on emergent surfaces? (Some depth of water over the prints)
This I don’t know. But then I wouldn’t know how to pick out apatosaur tracks from those of other diplodocids, and I bet no-one else does, either. Tracks are notoriously variable in shape, and can very wildly from the that of the feet that made them. Given that diplodocid feet were mostly pretty similar anyway, I would not be easily persuaded that any track can be confidently identified down to the genus level.
One other thing to be aware of is that there is often not agreement on the conditions under which a given track is made. One palaeontologist may think a given a track is a direct print, another will think it’s an underprint. I don’t mean to say that it’s hopeless and all we can do is throw our hands up in despair — good work is being done on interpreting tracks, but we have a long way to go. And this is not an area that I’m at all expert in.
Question #5
I read recently that in order for a skin impression to be made the Dermal tissue must undergo a type of chemical alteration! Do you think this is what allows the impression to be made?
That doesn’t sound right to me. The first thing that has to happen for an animal to be fossilized is that it needs to be buried in sediment really fast after it dies — before it’s eaten by scavengers. For something as fine as skin impressions to be preserved, that sediment needs to be very fine — which sadly tends to conflict with the first condition, since course sediments can be deposited more quickly than fine ones. It’s really hard for enough fine sediment to be laid down quickly enough to cover an animal of the size of a sauropod, which is why we don’t have sauropod specimens like those gloriously preserved theropods from the Yixian Formation in China(*). So sauropod skin impressions are pretty rare.
(*) Alternatively: there are spectacularly preserved partial sauropod specimens in the Yixian, but Chinese researchers can’t be bothered to write them up because they’d rather spend their time getting a slam-dunk Nature paper out of yet another little feathered theropod. Unduly cynical? Maybe. But I continue to live in hope.
Well, that about wraps up the five questions — to the best of my ability at least. But I’d love to hear from people who know more than I about these topics: leave a comment, and fame and glory could be yours!
And finally … if you, too, would like to have us answer five questions on the sauropod-related topic of your choice, quite possibly in the less than three years, you should consider getting yourself across the The SV-POW! Patreon and making an unreasonably extravagant financial commitment.
References
- Berman David S., and Bruce M. Rothschild. 2005. Neck posture of sauropods determined using radiological imaging to reveal three-dimensional structure of cervical vertebrae. pp. 233–247 in: Virgina Tidwell and Ken Carpenter, Thunder-lizards: the sauropodomorph dinosaurs. Bloomington: Indiana University Press.
- Taylor, Michael P. 2014. Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs. PeerJ 2:e712. doi: 10.7717/peerj.712
- Taylor, Michael P., David W. E. Hone, Mathew J. Wedel and Darren Naish. 2011. The long necks of sauropods did not evolve primarily through sexual selection. Journal of Zoology 285:150-161. doi: 10.1111/j.1469-7998.2011.00824.x
- Taylor, Michael P., Mathew J. Wedel, Darren Naish and Brian Engh. 2015. Were the necks of Apatosaurus and Brontosaurus adapted for combat?. p. 71 in Mark Young (ed.), Abstracts, 63rd Symposium for Vertebrate Palaeontology and Comparative Anatomy, Southampton. 115 pp.
If I could dissect a sauropod…
September 12, 2024
Luke Horton asked in a comment on a recent post:
Given the chance to examine a titanosaur cadaver with your hypothetical army of anatomists, what would you look for first?
*FACEPALM* How we’ve gone almost 17 years without posting about a hypothetical sauropod dissection is quite beyond my capacity. I am also contractually obligated to remind you that the TV show “Inside Nature’s Giants” shows dissections of a whale, elephant, giraffe, tiger, anaconda, giant squid, etc., so it’s probably the closest we’ll ever get. Go look up photos of Dr. Joy Reidenberg standing, um, amidst a partially-dissected whale, or just watch that episode, and your sauropod-dissection-visualizer will be properly calibrated.
To get back to Luke’s question, there are loads of interesting things that could be dissected in a sauropod, but since the remit here is Matt Wedel x titanosaur, there’s only one possible answer: the lung/air sac system and its diverticula. For several reasons:

Hypothetical reconstruction of the lungs (red) and air sacs (blue, green, and gray) in Haplocanthosaurus CM 879. I’d love to know how close this is to reality. Wedel (2009: fig. 10).
First and most obviously, I’ve spent the last quarter-century trying to infer as much as possible about the respiratory systems of sauropods based on the patterns of pneumaticity in their skeletons, and I’d kill for the opportunity to check the accuracy of my inferences — and those of all my fellow-travelers in the sauropod and dinosaur respiration biz, like Daniela Schwarz and Emma Schachner and Tito Aureliano and many others.

Sauropod respiratory system modeled on that of a bird. I’ll bet the correspondence wasn’t this close. (Also, since making this figure 20 years ago, I’ve learned that the abdominal air sacs of ostriches are actually rather small, although the perirenal, femoral, and subcutaneous diverticula of the abdominal air sacs are extensive; see Bezuidenhout et al. 1999). Wedel and Cifelli (2005: fig. 14).
Second, I am intrigued/haunted by the possibility that extant birds might not represent the apex of saurischian lung/air sac evolution. Birds survived the K-Pg disaster because they were small; respiratory efficiency had little or nothing to do with it (evidence: all the other small-bodied tetrapods that survived, like the many, many squamate and mammalian lineages). To me it would be a wild coincidence if the tiny dinosaurs that survived also just happened to be The Bestest (TM) at some anatomical/physiological thing unrelated to their survival. In fact, given how sensitive birds are to airborne dust and ash, I wonder if their fancy lungs weren’t more of a hindrance than a help in the dusty, sooty, iridium-laced post-impact world. Anyway, there are interesting clues that the air sac systems of extant birds are just one subset of a much greater original diversity, like most (all?) birds starting out embryologically with a dozen or so air sacs, which get simplified to the usual 9 or fewer by fusions. What did other dinosaurs do with their 12 (or more?) air sacs? If any dinosaurian clade was going to push the capabilities of the “avian” lung/air sac system in interesting directions and to fascinating extremes, sauropods seem like a good bet.

Rib articulation angles in the dorsal vertebrae of (a) Lufengosaurus, (b) Diplodocus, (c) Haplocanthosaurus, (d) Tyrannosaurus, and (e) an ostrich. Anterior is to the right. Diplodocus and Haplocanthosaurus are pretty wildly different considering they coexisted in the Morrison. I really gotta write a whole post about that. Boisvert et al. (2024: fig. 12).
So I’m intrigued by the idea that extant birds show us one way that a saurischian lung/air sac system can work, but don’t exhaust the territory, anymore than kangaroos show us all the ways that mammals can reproduce. Maybe sauropods had even better lungs than birds! Or maybe not. Likely they were doing their own weirdly specialized thing — or many weirdly specialized things — that left few to no diagnostic traces in their skeletons. We can be pretty confident that at least some of the pneumatic diverticula of sauropods worked essentially identically to how they do in birds (see Woodruff et al. 2022 and this post), and mid-dorsal pneumatic hiatuses in juvenile sauropods — predicted by me in 2003, found by Melstrom et al. (2016) and Hanik et al. (2017) — suggest that their air sac systems were broadly comparable. On the other hand, the variety of rib articulation angles just within Morrison sauropods tells us they weren’t all ventilating their air sacs in quite the same way (Boisvert et al. 2024), despite broad similarities with other dinos at the levels of rib osteology (Wang et al. 2023) and whole-thorax construction (Schachner et al. 2009, 2011). (Aside: why the hell didn’t I work a citation of Wang et al. 2023 into the Dry Mesa Haplo paper? I can only conclude that I am at least occasionally an idiot.) Whatever was going on, I’m pretty sure sauropods didn’t look exactly like 60-ton turkeys on the inside, but we don’t have a ton of real data on how they differed. It would be amazing to find out.

The mounted Rapetosaurus skeleton at the Field Museum, traced from a photo. Specific weird things to note: neck about twice as long as tail, cervical vertebrae about twice as tall as dorsals, and smallish pelvic bones relative to hindlimbs (= skinny posterior abdomen, at least dorsoventrally). See this post for details.
Third, if any sauropods were going to rival or exceed birds in fancy under-the-hood anatomical and physiological adaptations, my money would be on titanosaurs. They were morphologically disparate, phylogenetically diverse, geographically widespread, they independently evolved to giant size more times than any other sauropod clade, and their growth rates were wild. I’d dissect any sauropod I got access to (uh duh), but a titanosaur would be particularly appealing. Which titanosaur? Probably Rapetosaurus: we know it grew very fast early on (Curry Rogers et al. 2016, and see implications for the nervous system in Smith et al. 2022), it had a highly pneumatic vertebral column (O’Connor 2006), its body proportions were pretty wacky, and it had other features of interest to me, like expanded neurocentral joints (see Wedel and Atterholt 2023 and this post) and neural canal ridges (see Atterholt et al. 2024 and this post).

I used this photo of a Rapetosaurus caudal vertebra a few posts ago to illustrate the neural canal ridges, but — like many other sauropods — it also has very expanded neurocentral joints forming boutons. From Curry Rogers (2009: fig. 27).
Oh, and if I got to dissect more than one sauropod, the rest of my top 5 choices in order would be:
- the owner of BYU 9024 (Supersaurus? Giant ancient individual of Barosaurus? Are those even different things? Dissecting this critter could tell us!), Barosaurus being the most diplodocid-y and least titanosaur-y neosauropod I know of, and BYU 9024 being from a hellaciously big individual no matter what its classification;
- the Snowmass Haplocanthosaurus, because I have just so many questions about all the weird stuff going on with its tail (see Wedel et al. 2021 and this post for starters);
- Omeisaurus or Xinjiangtitan, to represent a maximally derived-but-also-weird non-neosauropod;
- Sauroposeidon, for obvious emotional reasons (but not enough to dethrone the others).
After that? Probably Isanosaurus or Melanorosaurus or something else waaaay down the tree, so I could see how much of the sauropod kit was in place from the get-go (probably most of it).

Bone vs joint space in the proximal caudals of the Snowmass Haplocanthosaurus. I’d give one non-essential organ to dissect that tail!
And after the respiratory system, next up for me would be the spinal cord and any related morphological specializations of the neural canal — see Table 3 in Atterholt et al. (2024) for a running tally, and this page. Then intervertebral joints, digestive tract, and reproductive system (neither of the last two leave anything useful in the way of skeletal traces), in that order. Arguably the intervertebral joints would be a bigger score for sauropod paleobiology than spinal cord stuff, but maybe not, and having squelched my emotional pick among sauropod taxa, I’m letting my emotions rule when choosing body systems to dissect. I also am intensely interested in the possibility of protofeathers in sauropods, but you don’t have to dissect those, you can just see if any are present, so I’d cheat a little and note any integumentary specializations en passant. (Remember than an animal can have hairs without being hairy [naked mole rats, rhinos, manatees, dolphins], ditto for feathers.)
So that’s the sauropod and the body system I’d dissect first, if given the chance. What’s your answer?
References
- Atterholt, J., Wedel, M.J., Tykoski, R., Fiorillo, A.R., Holwerda, F., Nalley, T.K., Lepore, T., and Yasmer, J. 2024. Neural canal ridges: a novel osteological correlate of postcranial neuroanatomy in dinosaurs. The Anatomical Record, 1-20. https://doi.org/10.1002/ar.25558
- Bezuidenhout, A.J., H.B. Groenewald, and J.T. Soley. 1999. An anatomical study of the respiratory air sacs in ostriches. Onderstepoort Journal of Veterinary Research 66:317-325.
- Boisvert, Colin, Curtice, Brian, Wedel, Mathew, & Wilhite, Ray. 2024. Description of a new specimen of Haplocanthosaurus from the Dry Mesa Dinosaur Quarry. The Anatomical Record, 1–19. http://doi.org/10.1002/ar.25520
- Curry Rogers, Kristina. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 29:1046-1086.
- Curry Rogers, K., M. Whitney, M. D. D’Emic, and B. Bagley. 2016. Precocity in a tiny titanosaur from the Late Cretaceous of Madagascar. Science 352:450–454.
- Hanik, Gina M., Matthew C. Lamanna and John A. Whitlock. 2017. A juvenile specimen of Barosaurus Marsh, 1890 (Sauropoda: Diplodocidae) from the Upper Jurassic Morrison Formation of Dinosaur National Monument, Utah, USA. Annals of Carnegie Museum 84(3):253–263.
- Melstrom, Keegan M., Michael D. D’Emic, Daniel Chure and Jeffrey A. Wilson. 2016. A juvenile sauropod dinosaur from the Late Jurassic of Utah, USA, presents further evidence of an avian style air-sac system. Journal of Vertebrate Paleontology 36(4):e1111898. doi:10.1080/02724634.2016.1111898
- O’Connor, P.M. 2006. Postcranial pneumaticity: an evaluation of soft-tissue influences on the postcranial skeleton and the reconstruction of pulmonary anatomy in archosaurs. Journal of Morphology 267: 1199-1226.
- Schachner, E.R., Lyson, T.R. and Dodson, P., 2009. Evolution of the respiratory system in nonavian theropods: evidence from rib and vertebral morphology. The Anatomical Record 292(9): 1501-1513.
- Schachner, E.R., Farmer, C.G., McDonald, A.T. and Dodson, P., 2011. Evolution of the dinosauriform respiratory apparatus: new evidence from the postcranial axial skeleton. The Anatomical Record 294(9): 1532-1547.
- Smith, Douglas H., Rodgers, Jeffrey M., Dollé, Jean-Pierre, and Wedel, Mathew J. 2022. Giraffes vs. blue whales vs. dinosaurs: contest reveals which one builds its nervous system fastest to evade predators. Scientific American, https://www.scientificamerican.com/article/giraffes-vs-blue-whales-vs-dinosaurs-contest-reveals-which-one-builds-its-nervous-system-fastest-to-evade-predators/
- Wang, Y.Y., Claessens, L.P. and Sullivan, C., 2023. Deep reptilian evolutionary roots of a major avian respiratory adaptation. Communications Biology, 6(1), p.3.
- Wedel, M.J. 2003a. Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. Paleobiology 29:243-255.
- Wedel, M.J. 2009. Evidence for bird-like air sacs in saurischian dinosaurs. Journal of Experimental Zoology 311A:611-628.
- Wedel, M.J., and Atterholt, J. 2023. Expanded neurocentral joints in the vertebrae of sauropod dinosaurs. In Hunt-Foster, R.K., Kirkland, J.I., and Loewen, M.A. (eds), 14th Symposium on Mesozoic Terrestrial Ecosystems and Biota. The Anatomical Record 306(S1):256-257.
- Wedel, M.J., and Cifelli, R.L. 2005. Sauroposeidon: Oklahoma’s native giant. Oklahoma Geology Notes 65 (2):40-57.
- Wedel, Mathew; Atterholt, Jessie; Dooley, Jr., Alton C.; Farooq, Saad; Macalino, Jeff; Nalley, Thierra K.; Wisser, Gary; and Yasmer, John. 2021. Expanded neural canals in the caudal vertebrae of a specimen of Haplocanthosaurus. Academia Letters, Article 911, 10pp.
- Woodruff, D. Cary, Wolff, Ewan D.S., Wedel, Mathew J., Dennison, Sophie, and Witmer, Lawrence M. 2022. The first occurrence of an avian-style respiratory infection in a non-avian dinosaur. Scientific Reports 12, 1954. https://doi.org/10.1038/s41598-022-05761-3
Over on Mastodon (sign up, it’s great!), Jim Kirkland posted a baby Utahraptor caudal vertebrae for #FossilFriday. Here it is:

And after a bit of virtual prep work:

My first reaction was just “That’s pretty!“. My second, which I admit should have been my first, was “Wait a sec — how the heck do those things articulate?”
The issue is that both the prezygs and the postzygs overhang the centrum by so much. If we imagine three of these babies consecutively, there are basically two options.
First, the centra articulate closely, with what we might feel intuitively is a reasonable cartilage gap; and the zygs cross over:

Does something like this ever happen? Not in sauropods, for sure, but it could be correct — if the zyg facets are some way short of the tips of their processes, so that the most distal parts of each process are pre-epipophyses and epipophyses rather than prezygs and postzygs per se.
The other interpretation is this, with the zygs overlapping near the end as in sensible dinosaurs, and much more spaced out centra:

If this is right, then (in this respect) baby Utahraptor tails resembled camel necks in having big intervertebral spaces, which in life were filled with big cartilage plugs.
.
.
.
SPOILER SPACE
.
.
.
Have a think about this before reading on.
.
.
.
SPOILER SPACE
.
.
.
OK, here is the horrible truth.
Dromaeosaur tails do overlap their zyg processes as in the first mock-up above: but they do much, much worse than this!
Here is the truly perverted figure 37 of Ostrom’s classic 1969 monograph on Deinonychus — the publication that catalysed the whole Dinosaur Renaissance:

As you can see, the zygapophyseal processes are grotesquely elongated, and overlap in long stiffening bundles with those of successive vertebrae (part C of the figure). The actual zyg facets are small, and close to the origins of these processes (see parts A and B of the figure). And the chevrons are also hideously protracted beyond their natural length to form stiffening bundles beneath the tail that complement those above the tail.
To add insult to injury, the chevrons even face in the wrong darned direction, extending anteriorly along the tail rather than posteriorly as in all decent animals. Yes: in Ostrom’s illustrations, we’re seeing the vertebrae in right lateral view, i.e. anterior is to the right.
All of this confirms that I was so, so right two decades ago to focus so completely on proper dinosaurs instead of these nasty mutant ones. Ugh.
What’s up with your insanely thick intervertebral discs, Snowmass Haplocanthosaurus?
February 28, 2022
Among the numerous weird features of MWC 8028, the Snowmass Haplocanthosaurus, is the extreme biconcave profile of the caudal vertebrae, in which each centrum is basically reduced to a vertical plate of bone separating two cup-shaped articular surfaces. All four available caudals — found in different parts of the quarry, in different orientations — have essentially the same cross-section. For the diagram above, I just copied caudal 3, because it’s the most complete, so I could figure out the thickness and cross-sectional shape of a single intervertebral disc.
I drew a more realistic version, with the first three caudals at approximately the right scale, for our neural canal paper last year:

The first three caudal vertebrae of Haplocanthosaurus specimen MWC 8028 in midsagittal section, emphasizing the volumes of the neural canal (yellow) and intervertebral joint spaces (blue). Anterior is to the right. Wedel et al. (2021: fig. 2B).
It’s a drawing, sure, but it’s based on a true story, because we have CT scans of all the vertebrae (and we’re going to publish them, soon, along with the reconstructed verts).
(NB: I’m using “intervertebral disc” as a convenient shorthand for “whatever soft tissues filled the joint space”. But I do think it was a big, fat, fibrocartilaginous disc, not wildly different from the ones in the human vertebral column. It’s not totally impossible that there was some combination of crazy thick articular cartilage and a synovial cavity — there is some precedent in extant salamanders and lizards — but that seems way less likely, for reasons I’ll go into in detail elsewhere. Incidentally, the notion is floating around that reptiles have only synovial intervertebral joints, but this is simply false: intervertebral discs are present in some squamates [Winchester and Bellairs 1977] and in the tails of birds [Baumel 1988].)
I should point out that the other specimens of Haplocanthosaurus also have biconcave caudal vertebrae, but the concavities are much shallower. So what we’re seeing in MWC 8028 is an extreme version of something we see in other individuals of the same genus.
Now, because the caudal centra and joint spaces are roughly radially symmetrical, their relative cross-sectional areas, in these mid-sagittal sections, should be good proxies for their relative volumes. You can imagine the generating the volume of a centrum by rotating its cross-section through 180 degrees, ditto for the joint space (ignoring tilt since both the centrum and joint space are tilted). We’ll have this math worked out in more detail in the next paper, along with volumes from the 3D models, but the upshot is this:
The volume of the intervertebral discs is about twice that of the vertebral centra. If we ignore the neural arch and spine and the transverse processes, and focus only on the weight-bearing column formed by the proximal caudal centra and intervertebral discs, that column is 2/3 cartilage and only 1/3 bone.
Why, tho?
I spent some time brainstorming with Alton Dooley and we came up with a whole slate of hypotheses. We don’t necessarily like any of them very much, we’re just trying to cast the widest possible net, to make sure we haven’t overlooked any possibilities, no matter how remote they might seem. Here’s what we have so far:
Non-biological:
1. taphonomic distortion
Abnormal biology:
2. congenital malformation
3. pathology
Ontogenetic:
4. incomplete ossification (animal died without laying down the ‘missing’ bone)
5. senescence (the ‘missing’ bone was removed by some process related to aging)
Functional:
6. increased or decreased movement between vertebrae
7. weight reduction
8. shock absorption
What else?
To reiterate, we’re in the hypothesis-generating stage, not the hypothesis-evaluating stage. So we’re not interested in whether any of these hypotheses are likely. (In point of fact, I think the ones we have so far all suck.) We just want all of the ideas that aren’t impossible.
The comment field is open!
References
- Baumel, J.J. 1988. Functional morphology of the tail apparatus of the pigeon (Columba livia). Advances in Anatomy, Embryology, and Cell Biology 110: 1-115.
- Wedel, Mathew; Atterholt, Jessie; Dooley, Jr., Alton C.; Farooq, Saad; Macalino, Jeff; Nalley, Thierra K.; Wisser, Gary; and Yasmer, John. 2021. Expanded neural canals in the caudal vertebrae of a specimen of Haplocanthosaurus. Academia Letters, Article 911, 10pp.
- Winchester, L. and Bellairs, A.D.A. 1977. Aspects of vertebral development in lizards and snakes. Journal of Zoology 181(4): 495-525.
What can sauropod sacra tell us about neck posture?
April 22, 2020
Daniel Vidal et al.’s new paper in Scientific Reports (Vidal et al. 2020) has been out for a couple of days now. Dealing as it does with sauropod neck posture, it’s obviously of interest to me, and to Matt. (See our earlier relevant papers Taylor et al. 2009, Taylor and Wedel 2013 and Taylor 2014.)

Overview
To brutally over-summarise Vidal et al.’s paper, it comes down to this: they digitized the beautifully preserved and nearly complete skeleton of Spinophorosaurus, and digitally articulated the scans of the bones to make a virtual skeletal mount. In doing this, they were careful to consider the neutral pose of consecutive vertebrae in isolation, looking at only one pair at a time, so as to avoid any unconscious biases as to how the articulated column “should” look.
Then they took the resulting pose, objectively arrived at — shown above in their figure 1 — and looked to see what it told them. And as you can well see, it showed a dramatically different pose from that of the original reconstruction.

Original skeletal reconstruction of Spinophorosaurus nigerensis (Remes et al. 2009:figure 5, reversed for ease of comparison). Dimensions are based on GCP-CV-4229/NMB-1699-R, elements that are not represented are shaded. Scale bar = 1 m.
In particular, they found that as the sacrum is distinctly “wedged” (i.e. its anteroposterior length is greater ventrally than it is dorsally, giving it a functionally trapezoidal shape, shown in their figure 1A), so that the column of the torso is inclined 20 degrees dorsally relative to that of the tail. They also found lesser but still significant wedging in the last two dorsal vertebrae (figure 1B) and apparently some slight wedging in the first dorsal (figure 1C) and last cervical (figure 1D).
The upshot of all this is that their new reconstruction of Spinophorosaurus has a strongly inclined dorsal column, and consequently a strongly inclined cervical column in neutral pose.
Vidal et al. also note that all eusauropods have wedged sacra to a greater or lesser extent, and conclude that to varying degrees all eusauropods had a more inclined torso and neck than we have been used to reconstructing them with.
Response
I have to be careful about this paper, because its results flatter my preconceptions. I have always been a raised-neck advocate, and there is a temptation to leap onto any paper that reaches the same conclusion and see it as corroboration of my position.
The first thing to say is that the core observation is absolutely right, — and it’s one of those things that once it’s pointed out it’s so obvious that you wonder why you never made anything of it yourself. Yes, it’s true that sauropod sacra are wedged. It’s often difficult to see in lateral view because the ilia are usually fused to the sacral ribs, but when you see them in three dimensions it’s obvious. Occasionally you find a sacrum without its ilium, and then the wedging can hardly be missed … yet somehow, we’ve all been missing its implications for a century and a half.
Sacrum of Diplodocus AMNH 516 in left lateral and (for our purposes irrelevant) ventral views. (Osborn 1904 figure 3)
Of course this means that, other thing being equal, the tail and torso will not be parallel with each other, but will project in such a way that the angle between them, measured dorsally, is less than 180 degrees. And to be fair, Greg Paul has long been illustrating diplodocids with an upward kink to the tail, and some other palaeoartists have picked up on this — notably Scott Hartman with his very uncomfortable-looking Mamenchisaurus.
But I do have three important caveats that mean I can’t just take the conclusions of the Vidal et al. paper at face value.
1. Intervertebral cartilage
I know that we have rather banged on about this (Taylor and Wedel 2013, Taylor 2014) but it remains true that bones alone can tell us almost nothing about how vertebrae articulated. Unless we incorporate intervertebral cartilage into our models, they can only mislead us. To their credit, Vidal et al. are aware of this — though you wouldn’t know it from the actual paper, whose single mention of cartilage is in respect of a hypothesised cartilaginous suprascapula. But buried away the supplementary information is this rather despairing paragraph:
Cartilaginous Neutral Pose (CNP): the term was coined by Taylor for “the pose found when intervertebral cartilage [that separates the centra of adjacent vertebrae] is included”. Since the amount of inter-vertebral space cannot be certainly known for most fossil vertebrate taxa, true CNP will likely remain unknown for most taxa or always based on estimates.
Now this is true, so far as it goes: it’s usually impossible to know how much cartilage there was, and what shape it took, as only very unusual preservational conditions give us this information. But I don’t think that lets us out from the duty of recognising how crucial that cartilage is. It’s not enough just to say “It’s too hard to measure” and assume it didn’t exist. We need to be saying “Here are the results if we assume zero-thickness cartilage, here’s what we get if we assume cartilage thickness equal to 5% centrum length, and here’s what we get if we assume 10%”.
I really don’t think it’s good enough in 2020 to say “We know there was some intervertebral cartilage, but since we don’t know exactly how much we’re going to assume there was none at all”.
The thing about incorporating cartilage into articulating models is that we would, quite possibly, get crazy results. I refer you to the disturbing figure 4 in my 2014 paper:

Figure 4. Effect of adding cartilage to the neutral pose of the neck of Diplodocus carnegii CM 84. Images of vertebra from Hatcher (1901:plate III). At the bottom, the vertebrae are composed in a horizontal posture. Superimposed, the same vertebrae are shown inclined by the additional extension angles indicated in Table 2.
I imagine that taking cartilage into account for the Spinophorosaurus reconstruction might have given rise to equally crazy “neutral” postures. I can see why Vidal et al. might have been reluctant to open that can of worms; but the thing is, it’s a can that really needs opening.
2. Sacrum orientation
As Vidal et al.’s figure 1A clearly shows, the sacrum of Spinophorosaurus is indeed wedge-shaped, with the anterior articular surface of the first sacral forming an angle of 20 degrees relative to the posterior articular surface of the last:
But I don’t see why it follows that “the coalesced sacrum is situated so that the posterior face of the last sacral centrum is sub-vertical. This makes the presacral series slope dorsally and allows the tail to be subhorizontal (Figs. 1 and 4S)”. Vidal et al. justify this by saying:
Since a subhorizontal tail has been known to be present in the majority of known sauropods[27, 28, 29], the [osteologically induced curvature] of the tail of Spinophorosaurus is therefore compatible with this condition.
But those three numbered references are to Gilmore 1932, Coombs 1975 and Bakker 1968 — three venerable papers, all over fifty years old, dating from a period long before the current understanding of sauropod posture. What’s more, each of those three was about disproving the previously widespread assumption of tail-dragging in sauropods, but the wedged sacrum of Spinophorosaurus if anything suggests the opposite posture.
So my question is, given that the dorsal and caudal portions of the vertebral column are at some specific angle to each other, how do we decide which (if either) is horizontal, and which is inclined?

Three interpretations of the wedged sacrum of Spinophorosaurus, in right lateral view. In all three, the green line represents the trajectory of the dorsal column in the torso, and the red line that of the caudal column. At the top, the tail is horizontal (as favoured by Vidal et al. 2020) resulting in an inclined torso; at the bottom, the torso is horizontal, resulting in a dorsally inclined tail; in the middle, an intermediate posture shows both the torso and the tail slightly inclined.
I am not convinced that the evidence presented by Vidal et al. persuasively favours any of these possibilities over the others. (They restore the forequarters of Spinophorosaurus with a very vertical and ventrally positioned scapula in order to enable the forefeet to reach the ground; this may be correct or it may not, but it’s by no means certain — especially as the humeri are cross-scaled from a referred specimen and the radius, ulna and manus completely unknown.)
3. Distortion
Finally, we should mention the problem of distortion. This is not really a criticism of the paper, just a warning that sacra as preserved should not be taken as gospel. I have no statistics or even systematic observations to back up this assertion, but the impression I have, from having looked closely at quite a lot of sauropod vertebra, is the sacra are perhaps more prone to distortion than most vertebrae. So, for example, the very extreme almost 30-degree wedging that Vidal et al. observed in the sacrum of the Brachiosaurus altithorax holotype FMNH PR 25107 should perhaps not be taken at face value.
Now what?
Vidal el al. are obviously onto something. Sauropod sacra are screwy, and I’m glad they have drawn attention in a systematic way to something that had only been alluded to in passing previously, and often in a way that made it seems as though the wedging they describe was unique to a few special specimens. So it’s good that this paper is out there.
But we really do need to see it as only a beginning. Some of the things I want to see:
- Taking cartilage into account. If this results in silly postures, we need to understand why that is the case, not just pretend the problem doesn’t exist.
- Comparison of sauropod sacra with those of other animals — most important, extant animals whose actual posture we can observe. This might be able to tell us whether wedging really has the implications for posture that we’re assuming.
- Better justification of the claim that the torso rather than the tail was inclined.
- An emerging consensus on sauropod shoulder articulation, since this also bears on torso orientation. (I don’t really have a position on this, but I think Matt does.)
- The digital Spinophorosaurus model used in this study. (The paper says “The digital fossils used to build the virtual skeleton are deposited and accessioned at the Museo Paleontológico de Elche” but there is no link, I can’t easily find them on the website and they really should be published alongside the paper.)
Anyway, this is a good beginning. Onward and upward!
References
- Bakker, Robert T. 1968. The Superiority of Dinosaurs. Discovery 3:11–22.
- Coombs, Walter P. 1975. Sauropod habits and habitats. Palaeogeography, Palaeoclimatology, Palaeoecology 17:1-33.
- Gilmore, Charles W. 1932. On a newly mounted skeleton of Diplodocus in the United States National Museum. Proceedings of the United States National Museum 81:1-21.
- Hatcher, John Bell. 1901. Diplodocus (Marsh): its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1:1-63.
- Osborn, Henry F. 1904. Manus, sacrum and caudals of Sauropoda. Bulletin of the American Museum of Natural History 20:181-190.
- Taylor, Michael P. 2014. Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs. PeerJ 2:e712. doi:10.7717/peerj.712
- Taylor, Michael P., and Mathew J. Wedel. 2013c. The effect of intervertebral cartilage on neutral posture and range of motion in the necks of sauropod dinosaurs. PLOS ONE 8(10):e78214. 17 pages. doi:10.1371/journal.pone.0078214
- Taylor, Michael P., Mathew J. Wedel and Darren Naish. 2009. Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54(2):213-230.
- Vidal, Daniel, P Mocho, A. Aberasturi, J. L. Sanz and F. Ortega. 2020. High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs. Scientific Reports 10(6638). Indispensible supplementary information at https://static-content.springer.com/esm/art%3A10.1038%2Fs41598-020-63439-0/MediaObjects/41598_2020_63439_MOESM1_ESM.pdf
doi:10.1038/s41598-020-63439-0
The Haplocanthosaurus presentation from the 1st Palaeo Virtual Congress is now a PeerJ Preprint
December 17, 2018

If you were curious about the Wedel et al. presentation on the Snowmass Haplocanthosaurus at the 1st Palaeo Virtual Congress but didn’t attend the event, it is now preserved for posterity and freely available to the world as a PeerJ Preprint (as promised). Here’s the link.
I’ll have much more to say about this going forward, but for now here are slides 20 and 21 on the intervertebral joint spaces. This is obviously just the same vert cloned three times and articulated with itself. With the digital rearticulation of the reconstructed and retrodeformed caudal series still in progress, we cloned caudal 3, the only vertebra that preserves both sets of zygapophyses, to get a rough estimate of the sizes and shapes of the soft tissues that filled the intervertebral spaces and neural canal.

The reconstructed intervertebral discs (in blue) are very crude and diagrammatic. The reason I’m putting these particular slides up is to get the cited references out in the open on the blog, to start correcting the misapprehension that all non-mammalian amniotes have exclusively synovial intervertebral joints (see the discussion in the comments on this post). In the list below I’m including Banerji (1957), which is not cited in the presentation but which I did cite in that comment thread; it’s an important source and at least for now it is a free download. These refs are just the tip of a very big iceberg. One of my goals for 2019 is to do a series of posts reviewing the extensive literature on amphiarthrodial (fibrocartilaginous) intervertebral joints in living lepidosaurs and birds. Stay tuned!
And please go have a look at the presentation if you are at all interested or curious. As we said in the next to last slide, “this research is ongoing, and we welcome your input. If there are facts or hypotheses we haven’t considered but should, please let us know!”
References
- Baumel, J.J. (ed.) 1993. Handbook of Avian Anatomy: Nomina Anatomica Avium. Nuttal Ornithological Club, Publication No. 23, Cambridge, Massachusetts, USA.
- Banerji, K.K. 1957. The comparitive anatomy of the intervertebral joints. Doctoral dissertation, University of St Andrews, 132 + viii pp.
- Wedel, M.J., Atterholt, J., Macalino, J., Nalley, T., Wisser, G., and Yasmer, J. 2018. Reconstructing an unusual specimen of Haplocanthosaurus using a blend of physical and digital techniques. Abstract book, 1st Palaeontological Virtual Congress, http://palaeovc.uv.es/, p. 158 / PeerJ Preprints 6:e27431v1
- Winchester, L.A. 1978. Development and morphology of vertebrae and intervertebral joints in lizards and snakes. PhD thesis, Imperial College London, 326 pp.
- Winchester, L.A., and Bellairs, A.D.A. 1977. Aspects of vertebral development in lizards and snakes. J. Zoology 181(4): 495-525.
We may never know how flexible sauropod necks were
November 8, 2018
The more I look at the problem of how flexible sauropod necks were, the more I think we’re going to struggle to ever know their range of motion It’s just too dependent on soft tissue that doesn’t fossilise. Consider for example the difference between horse necks (above) and camel necks (below).


The skeletons of both consist of vertebrae that are pronouncedly opisthocoelous (convex in front and concave behind), so you might think their necks would be similarly flexible.
But the balls of horse cevicals are deeply embedded in their corresponding sockets, while those of camels have so much cartilage around and between them that the tip of the ball doesn’t even reach the rim of the socket. As a result of this (and maybe other factors), camel necks are far more flexible than those of horses.
Which do sauropod necks resemble? We don’t currently know, and we may never know. It will help if someone gets a good handle on osteological correlates of intervertebral cartilage.
[This post is recycled and expanded from a comment that I left on a Tetrapod Zoology post, but since Tet Zoo ate that comment it’s just as well I kept a copy.]
Sauropods’ neutral neck postures were really weird
November 5, 2014
Last night, I submitted a paper for publication — for the first time since April 2013. I’d almost forgotten what it felt like. But, because we’re living in the Shiny Digital Future, you don’t have to wait till it’s been through review and formal publication to read it. I submitted to PeerJ, and at the same time, made it available as a preprint (Taylor 2014).
It’s called “Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs”, and frankly the results are weird. Here’s a taste:
Taylor (2014:figure 3). Effect of adding cartilage to the neutral pose of the neck of Apatosaurus louisae CM 3018. Images of vertebra from Gilmore (1936:plate XXIV). At the bottom, the vertebrae are composed in a horizontal posture. Superimposed, the same vertebrae are shown inclined by the additional extension angles indicated in Table 1. If the slightly sub-horizontal osteological neutral pose of Stevens and Parrish (1999) is correct, then the cartilaginous neutral pose would be correspondingly slightly lower than depicted here, but still much closer to the elevated posture than to horizontal. (Note that the posture shown here would not have been the habitual posture in life: see discussion.)
A year back, as I was composing a blog-post about our neck-cartilage paper in PLOS ONE (Taylor and Wedel 2013c), I found myself writing down the rather trivial formula for the additional angle of extension at an intervertebral joint once the cartilage is taken into account. In that post, I finished with the promise “I guess that will have to go in a followup now”. Amazingly it’s taken me a year to get that one-pager written and submitted. (Although in the usual way of things, the manuscript ended up being 13 pages long.)
To summarise the main point of the paper: when you insert cartilage of thickness t between two vertebrae whose zygapophyses articulate at height h above the centra, the more anterior vertebra is forced upwards by t/h radians. Our best guess for how much cartilage is between the adjacent vertebrae in an Apatosaurus neck is about 10% of centrum length: the image above shows the effect of inserting that much cartilage at each joint.
And yes, it’s weird. But it’s where the data leads me, so I think it would be dishonest not to publish it.
I’ll be interested to see what the reviewers make of this. You are all of course welcome to leave comments on the preprint itself; but because this is going through conventional peer-review straight away (unlike our Barosaurus preprint), there’s no need to offer the kind of detailed and comprehensive comment that several people did with the previous one. Of course feel free if you wish, but I’m not depending on it.
References
Gilmore Charles W. 1936. Osteology of Apatosaurus, with special reference to specimens in the Carnegie Museum. Memoirs of the Carnegie Museum 11:175–300 and plates XXI–XXXIV.
Stevens, Kent A., and J. Michael Parrish. 1999. Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science 284(5415):798–800. doi:10.1126/science.284.5415.798
Does anyone want a project? How can we understand sauropod neck cartilage better?
September 27, 2014
A couple of times now, I’ve pitched in an abstract for a Masters project looking at neck cartilage, hoping someone at Bristol will work on it with me co-supervising, but so far no-one’s bitten. Here’s how I’ve been describing it:
Understanding posture and motion in the necks of sauropods: the crucial role of cartilage in intervertebral joints
The sauropod dinosaurs were an order of magnitude bigger than any other terrestrial animal. Much sauropod research has concentrated on their long necks, which were crucial to their success (e.g. Sander et al. 2010). One approach to understanding neck function tries to determine neutral posture and range of motion by modelling the cervical vertebrae as a mechanical system (e.g. Stevens and Parrish 1999).
The raw material of such studies is fossilised vertebrae, but these are problematic for several reasons. The invariable incompleteness and distortion of sauropod neck fossils causes fundamental difficulties; but even given perfect fossils, the lack of preserved cartilage means that the bones are not shaped or sized as they were in life.
Ignoring cartilage has dramatic consequences for neutral posture, range of motion and even length of necks: pilot studies (Cobley 2011, Taylor 2011) found that intact bird necks are 8–12% longer than articulated sequences of their dry bones, and that figure is as high as 24% for a juvenile giraffe neck. A turkey neck postzygapophysis was 26% longer when cartilage was included than after being stripped down to naked bone.
We do not yet know how much articular cartilage sauropods had in their necks, nor even what kind of intervertebral joints they had: crocodilians have fibrocartilaginous discs like those of mammals, while birds have synovial joints, so the extant phylogenetic bracket is uninformative.
The project will involve dissection and measurement of bird and crocodilian necks, documenting the extent and shape of articular cartilage, identifying osteological correlates of fibrocartilaginous and synovial joints, and applying this data to sauropods to determine the nature of their neck joints and length of their necks, to reconstruct the lost cartilage, and to determine its effect on neutral pose and range of motion.
Following completion, we anticipate publication of the project.
References
Cobley, Matthew J. 2011. The flexibility and musculature of the ostrich neck: implications for the feeding ecology and reconstruction of the Sauropoda (Dinosauria: Saurischia). MSc Thesis, Department of Earth Sciences, University of Bristol. vi+64 pages.
Sander, P. Martin, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva-Maria Griebeler, Hanns-Christian Gunga, Jürgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas Tütken, Oliver Wings and Ulrich Witzel. 2010. Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews 86:117–155. doi:10.1111/j.1469-185X.2010.00137.x
Stevens, Kent A., and J. Michael Parrish. 1999. Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs. Science 284:798–800. doi:10.1126/science.284.5415.798
Taylor, Michael P., and Mathew J. Wedel. 2011. Sauropod necks: how much do we really know?. p. 20 in Richard Forrest (ed.), Abstracts of Presentations, 59th Annual Symposium of Vertebrae Palaeontology and Comparative Anatomy, Lyme Regis, Dorset, UK, September 12th–17th 2011. 37 pp. http://www.miketaylor.org.uk/dino/pubs/svpca2011/TaylorWedel2011-what-do-we-really-know.ppt
(Obviously some part of this have since been covered by my and Matt’s first cartilage paper, but plenty has not.)
I now think there are two reasons no-one’s taken up this project: first, because I wrote it as very focussed only on the question of what type of joint was present, whereas there are plenty of related issues to be investigated along the way; and second, because I wrote it as a quest to discover a specific treasure (an osteological correlate), with the implication that if there’s no treasure to be found then the project will have been a failure.
But I do think there is still plenty of important work to be done in this area, and that there’s lots of important information to be got out of comparative dissection of extant critters.
If anyone out there fancies working in this area, I’d be delighted. I’d also be happy to offer whatever advice and help I could.
Update (18 October 2014)
Somehow I’d forgotten, when I wrote this post, that I’d previously written a more detailed post about the discs-in-sauropod-necks problem. If you’re interested in the problem, you should read that.

