Tutorial 48: my museum collections kit
November 26, 2025

I was on the road for most of August, September, and October, and in particular I made a ton of museum collections visits. When I visit a museum collection, I bring a specific set of gear that helps me get the photos, notes, and measurements that I want. All of this is YMMV — I’m not trying to predict what will work best for you, but to explain what has worked for me, and why. I’m reasonably happy with my current setup, but even after 28 years of museum visits, I’m still finding ways to improve it. Hence this post, which will hopefully serve as a vehicle for sharing tips and tricks.
A word about my program when I visit a collection, because not everyone needs or wants to do things my way. The closest museums with extensive sauropod collections are states away from where I live and work. If I’m in those collections at all, I’m traveling, and therefore on the clock. Time in collections is a zero-sum game: if I have the time to take 20 pages of notes, that could be 4 pages of notes of each of 5 specimens, 2 pages on 10, 1 page on 20, half a page on 40, etc. In practice, I usually make expansive notes early in the visit, one or two spreads per specimen with detailed sketches and exhaustive measurements of the most publication-worthy elements. I grade toward brevity over the course of the visit, and end with a mad desperate rush, throwing in crude sketches and rudimentary notes on as many newly-discovered (by me) specimens as possible. My collections visits are Discovery Time and Gathering Time, trying to get all the measurements and photographs I’ll want for the next year, or five, or forever. And, to the extent that I can suppress them, not Analysis Time or Graphing Time or Writing Time — I can do those things after hours and in my office back home, IF and only if I’ve spent my collections time efficiently gathering all the information I’ll need later.
The very first thing I do in any collection is a walking survey, to make sure I know roughly what specimens the collection contains and where to find them. For a sufficiently large collection — or even a single cabinet with 10 drawers of good stuff — I may draw a map in my notebook, on which I can note things I want to come back and document, and add new things as I find them.
Enough preamble, on to the gear. The first two or three entries here are in strict priority order, and after that things get very fuzzy and approximate.

1. Research Notebook
Seems obvious, right? Write stuff down, make sketches, capture the info that will be difficult or impossible to recapture later from photos. I have encountered people who don’t take a physical notebook, just a laptop or tablet, and take all their notes digitally. If that works for you, may a thousand gardens grow. For me, sketching is a fundamental activity — for fixing morphology in my mind, disciplining myself to see the whole object and its parts, creating a template on which to take further explanatory notes, and capturing the caveats, stray ideas, and odd connections that surround each specimen in a quantum fuzz in my mind (temporarily in my mind, hence the need for external capture). I also write priority lists in advance of specimens to document each day, and then cross them off, add new ones, and strike out duds with wild abandon in the heat of data collection.

I do a few specific things to increase the usefulness of my notebooks:
– Label the spines and covers with the notebook titles and years. These things live on the shelf directly over my desk, and I pull them down and rifle through them constantly. I also have notebooks for university service (committees, student advising, and so on), astronomical observations, and personal journaling, so “Research” is a useful tag for me.
– Number the pages, if they’re not already numbered, use the books chronologically from front to back, and create the table of contents retrospectively as I go — a tip I got from the Bullet Journal method.

– Paste a small envelope inside the back cover, if a pouch is not already built in, to hold all kinds of ephemera — index cards, scale bars, a bandage (just in case), stickers I acquire along the way, etc.

– Affix a section of measuring tape to the outer edge of the front or back cover. I got this tip from the naturalist John Muir Laws, whose Laws Guide to Nature Drawing and Journaling is wonderfully useful and inspiring (UPDATE: that book is now covered in its own post, here). The scale-bar-permanently-affixed-to-research-notebook has been a game-changer for me. Do you know how many times I’ve accidentally left a scale bar on a museum shelf, and then gotten to my next stop and had to borrow or fabricate one? I myself lost count long ago. But never again. If I’m in a hurry, small specimens go straight onto the notebook to be photographed, like the baby apatosaurine tibia above, and the notebook itself goes into the frame with large specimens. (This comes up again — if possible, and it’s almost always possible, put the specimen label in the photo with the specimen. No reason not to, and sometimes a lifesaver later on.)

Behold the thinness of the eminently pocketable IKEA paper tape. Folding instructions, because this seems to bedevil some folks: hold up one end, fold in half by grabbing the other end and bring it up in front, then do that three more times. Finished product is 65mm long, 25.4mm wide, and about 1mm thick when folded crisply and left under a heavy book overnight.
2. Measuring tapes
I find the flexible kind much more convenient and useful than retractable metal tape measures. I like the 1-2mm thick plastic type used by tailors and fabric sellers, because they have just enough inertia to stay where I put them, or drop in a predictable fashion when draped over something sufficiently large, as when measuring midshaft circumference of a long bone.
I LOVE the little plasticized paper tapes that hang on racks, free for the taking, near the entrances of IKEA stores. I tear them off by the dozen when I go to IKEA, cram them in my pockets, fold them flat when I get home, and stash them everywhere, including in my wallet. A few specific reasons they’re great:
– Folded flat, they’re about the thickness of a credit card, so there’s just no reason to be without one. I usually have one in my wallet, another in the envelope at the back of my research notebook, a couple more stashed in my luggage, a couple more stashed in my car, desk, tookbox, nightstand, etc.
– I can write on them. Especially handy if:
– I’ve torn off a section to serve as an impromptu scale bar. Which I never hesitate to do, because they’re free and I have dozens waiting in my toolbox and desk drawers at any one time. Torn off bits also make good bookmarks, classier, more cerebral, and less implicitly gross than the traditional folded square of toilet paper.
– I give them away to folks I’m traveling with, or that I meet in my travels, and they’re usually well-received.

I would NOT have figured out all these laminae if I hadn’t had a way to make them stand out.
3. Writing instruments in various colors
Up until about 2018 my notebooks were always monochrome pen or pencil. Then I realized that color is an extremely helpful differentiator for Future Matt, so now I highlight and color-annotate willy-nilly.
4. Calipers
I borrowed the digital calipers from Colin Boisvert to get the photo up top, having forgotten my own at home. As a sauropod worker, I don’t need sub-millimeter accuracy all the time. But digital calipers have three exceedingly useful functions: measuring the thickness of very thin laminae and bony septa; measuring the internal dimensions of small fossae and foramina; and measuring the depth of fossae and of concave articular surfaces. I also have a little titanium caliper on a lanyard that goes with me most places.
5. Small brush on a carabiner
This is the newest addition to the kit. I got the idea from Matthew Mossbrucker at the Morrison Museum in Morrison, Colorado. Colin and I visited him in September, immediately before our week-long stint in the collections at Dinosaur Journey. Matthew keeps a little brush carabinered to his belt at all times, and the utility was so instantly obvious that when Colin and I rolled into Fruita later that same day, I went to the hardware store and got my own. Cheap, weighs nothing, clips to anything, compact enough to cram in a pocket, good for lab and field alike. Genius!
6. Scale bar
Yes, I have my scale-bar-enhanced research notebook and my hoarder stash of IKEA paper tapes, but good old-fashioned scale bars are still useful, and I use them constantly. And lose them constantly, hence my multiple redundant backup mechanisms.
(Aside: I can’t explain why I hold onto some objects like grim death, but let others fall through my fingers like sand grains. I’ve only lost one notebook of any kind in my entire life — set it on top of the car while packing and then drove off [grrrr] — so I have no problem investing in nice notebooks and treating them like permanent fixtures. But I can’t hang onto pens and scale bars to save my life, hence my having gravitated to Bic sticks and IKEA paper tapes.)
7. Index cards
I try to get as much information into each photograph as possible. Ideally alongside the specimen I will have:
– a scale bar at the appropriate depth of field;
– the specimen tag with the number, locality, and other pertinent info;
– my notebook open to my sketch of the specimen, for easy correlation later (I don’t do this for every single view, just the ones that I think are particularly publication-worthy, or have info I’m likely to forget later);
– anything else I might want — serial position, anatomical directions, whether the photo is part of an anaglyph pair, and so on — written on an index card, which being a standard size will itself serve as an alternate/backup scale bar.
8. Pencil case
To hold all the smaller fiddly bits you see in the photo up top. I can’t now fathom why, but I resisted getting one of these for a loooong time. I was young and foolish then. Pretty useful all the time, absolutely clutch when it’s 4:58 pm and I’m throwing stuff in bags, caught between the Scylla of working as late as possible and the Charybdis of wanting to be polite to whatever kind, patient person is facilitating my visit. That is also when the pocket in the back of the notebook comes in especially handy.

Headlamp in action, casting low-angle light on a pneumatic fossa on the tuberculum of this sauropod rib. Note also the scale bar, elevated on a specimen box to be the same depth of field, and the notebook open to my sketch of the specimen.
9. Artificial lighting
This was another very late discovery for me — I don’t think I was regularly bringing my own lights prior to 2018. For me, portable, rechargeable lighting is useful in many circumstances and absolutely critical in two: casting low-angle light to pick out subtle pneumatic features, as in the photo above, and lighting up big specimens that I don’t have the time, energy, or space to pull off the shelves, as in the photo below.

I’m particularly taken with the big orange fan/light combo. It charges using a USB-C cable, has four settings for fan speed (handy when it’s hot, humid, or just oppressively still) and three for light intensity, a rotating hook that folds flat, and a USB power-out socket for charging phones, headlamps, fitness trackers, and what have you. I use it practically every day whether I’m on the road or not.

Magnetic flashlight hanging from steel shelving to illuminate Camarasaurus cervical vertebrae in the Utah Field House collections.
Whether it’s a hook or a magnet, some kind of mechanism for suspending a light at odd heights and angles is super useful. I usually have a strong flashlight with an integral seat-belt cutter and window-smasher in the door pocket of my car, and its magnetic base makes it omnidirectionally functional in collections spaces, which are usually liberally supplied with steel in the form of shelving and cabinets.

Haplocanthosaurus CM 879 caudal 2 in left lateral view, with rolled-up paper neural canal visualizer and scale-bar-stuck-to-flashlight.
Sometimes I use a bit of blue tack to stick a scale bar to a flashlight, to create a free-standing, truly vertical scale bar that I can rapidly place at different distances from the camera. Beats leaning the scale bar against a stack of empty specimen boxes or a block of ethofoam (which in turn beats nothing at all).
What else?
USUALLY — Laptop
Not for recording notes or measurements — all of that goes into the notebook, which I scan and upload new stuff from every evening. Mostly for displaying PDFs of descriptive monographs, and hugely useful in that regard.

MAYBE — Monographs
When I have the freedom (= baggage allowance) to do so, I find it handy to bring hardcopies of descriptive monographs, both for quick reference and so I can photograph specimens alongside the illustrations. Doesn’t even have to be the same specimens, just comparable elements. In the photo above, MWC 7257, a partial sacral centrum of Allosaurus from the Mygatt-Moore Quarry, is sitting next to a plate from Madsen (1976), illustrating the same vertebra in a specimen from Cleveland Lloyd Dinosaur Quarry. Thanks to Colin Boisvert for bringing the specimen to my attention — I’ve got a longstanding thing for sacrals — and for loaning me his copy of Madsen (1976) for this photo.
OUT — Camera and tripod
I suspect that some folks will shake their heads in mute horror, but after a couple of decades of lugging dedicated cameras and tripods everywhere, I stopped. For the past few years I’ve been rolling with just my phone, which is objectively better than any dedicated camera I owned for the first half of my career. Sometimes I brace it in an ad hoc fashion against a chair or shelf or cabinet, but mostly I just shoot freehand. For my purposes, it does fine, and any minor improvements in field curvature or whatever that I’d get from a dedicated camera don’t outweigh the logistical hassle. Again: YMMV!
Over to you
So, that’s what I roll with right now. It was different six months ago, and will almost certainly be a little different six months hence, hopefully as a result of people responding to this post. With all that said: what’s in your kit?
P.S. Many thanks to Matthew Mossbrucker and Julia McHugh for their hospitality and assistance in their collections, and to Colin Boisvert for being such a great travel companion, research sounding board, and generous loaner-of-things-I’d-forgotten. The Wedel-Boisvert Morrisonpocalypse 2025 deserves more blogging.

The pneumatic rib evolution figure in a more useful format
October 25, 2024
Here’s a funny thing I hadn’t given much thought to until recently: virtually all journals, even the born-digital variety, have pages in portrait mode for easy printing on 8.5×11 or A4 paper. And many offer a column-width option for figures. So if you want to line up a whole bunch of stuff for easy comparison, for a paper it’s usually easier to orient a figure vertically, like so:

Pneumatic dorsal ribs in a selection of sauropods and their outgroups. King et al. (2024: fig. 3).
And here it is in context on the page:

But virtually all slide presentations use a landscape format, 4:3 for a long time but often 16:9 these days to accommodate wider screens, or phones and tablets in landscape mode. For this a figure much taller than wide is usually not a good use of space, and may present at too small a scale to be readable.
I ran into this last week while prepping a presentation on my research for an anatomy department meeting at work. I wanted to use that King et al. figure because it summed up so much of the paper in one image, but the only version I had was the skyscraper version we’d used in the JVP paper. So I went into GIMP and rotated the image and every element within it by 90 degrees, to produce this landscape version:

I was presenting to an intellectually diverse audience, most of whom do not work on dinosaurs, so I added little silhouettes (my own, cribbed and hacked from all kinds of older work) to make it all more explicable:

This is all my original work, and I’m letting it out in the world here in case anyone else wants to use it. CC-BY like everything else on this blog. FWIW I think mamenchisaurs and diplodocids held their necks elevated — the baseline alert posture for extant tetrapods — I was just moving quickly and more concerned with getting little doodads for all the genera than with any paleobiological implications.
So now I’m wondering if there are any figures in old papers that I’ve avoided putting in talks, possibly subconsciously even, because they’re the wrong shape. Not that I need to do any more navel-gazing than I already do, but maybe something for me to keep an eye out for when I have reason to go back to them (which is often — they’re thought archives).
The more forward-looking takeaway is that if you have to make a taller-than-wide figure to fit a journal page, consider making a wider-than-tall version at the same time to throw into your talks — or vice versa if you’re making the talk first. It’s a time investment for sure, but it may be easier while all the bits are fresh in your head and you have all the elements in separate layers or whatever. Hopefully you already back up the uncompressed versions of all your figures, but Past Matt didn’t always do that, so at least be smarter than that guy!

Tate v2610, a sauropod dorsal rib. Check out the nice deep pneumatic fossa a little way down from the tuberculum of the rib (upper left in the photo).
Parting shot (and an excuse to post a photo for Fossil Friday): on my Tate trip this summer I hit a gang of museums, and everywhere I went I found pneumatic sauropod ribs. I think there are a lot more of these things out there than most folks have appreciated. I’m proud of my recent pneumatic rib papers (Taylor et al. 2023 and King et al. 2024), but I hope they are the just the start of something.
And because I picked that photo: you know what institution has a ton of super-interesting, well-preserved, well-prepped, not-yet-published-on sauropod vertebrae and ribs in a really nicely appointed collections room in an awesome museum run by a small team of excellent human beings? The Tate Geological Museum, that’s who. If you can get yourself to Casper and you have a legit research interest, go check out their collections, there’s SO MUCH good stuff in there. I myself will be back as soon as it can be conveniently arranged.
References
- King, J.L., McHugh, J.B., Wedel, M.J., and Curtice, B. 2024. A previously unreported form of dorsal rib pneumaticity in Apatosaurus (Dinosauria: Sauropoda) and implications for pneumatic variation among diplodocid dorsal ribs. Journal of Vertebrate Paleontology. DOI: 10.1080/02724634.2024.2316665
- Taylor, Michael P., and Matthew J. Wedel. 2023. Novel pneumatic features in the ribs of the sauropod dinosaur Brachiosaurus altithorax. Acta Palaeontologica Polonica 68(4): 709–718. doi:10.4202/app.01105.2023
Quick pix of ‘Jimbo’, the Wyoming Supersaurus
June 13, 2024

Another quick photo post from the road. The Tate Museum has a quality in common with the Oxford Museum of Natural History, where the guiding philosophy seems to have been, “Let’s put one of every interesting thing in the world in one big room.” Tucked into a corner is this small assemblage of cast bits of ‘Jimbo’, the Wyoming Supersaurus specimen described by Lovelace et al. (2008).

Here’s a tibia.

And a dorsal vertebra. I’m such a ninny, because the centrum is a little out-of-round I assumed that this was a cast of BYU 9044, the ‘Ultrasauros’ holotype vertebra. I didn’t figure out that it was a piece of Jimbo until I was on the road. *facepalm*
Anyway, in sauropod circles we refer to vertebrae like this as “real darn big”, the last size category before “stupidly huge”.

A dorsal rib, upside down. Pneumatic! Some cool art by Russell Hawley lurking behind.

And here’s the Jimbo mount at the Wyoming Dinosaur Center in Thermopolis.
Both the Tate and the WDC need a lot more nice things said about them by me, but this trip is still in progress, so all that will just have to wait.
Reference
I was cleaning out my Downloads directory — which, even after my initial forays, still accounts for 11 Gb that I really need to reclaim from my perptually almost-full SSD. And I found this beautiful image under the filename csgeo4028.jpeg.

Brachiosaurus altithorax holotype FMNH PR 25107 during excavation.
The thing is, I have no idea where this image came from. The file’s timestamp says it’s been 16 months since I downloaded it from somewhere, but there is no associated metadata that tells me where I got it. Googling for the filename gets me nothing.
Can anyone find the source? [Update: see the first coment below! Crown House found it in the Field Museum’s own Geology Collections.]
Anyway, I immediately recognised it as our old friend Brachiosaurus altithorax, and in fact it’s a much better version of a photo that we’ve featured here before, That version was scanned from Supplement 1 of Don Glut’s encyclopedia, which credits it as being Field Museum neg. #4027 (which is one out from the number in the filename). But that doesn’t explain where this high-resolution copy came from.
Anyway, looking at this image in 2024, I’m immediately interested in the ribs, which of course Matt and I published on at the very end of 2023 (Taylor and Wedel 2023, natch). It shows both ribs A and B in their original state, and it’s instructive to compare them with those ribs as we illustrated them in our paper.
First, rib A:
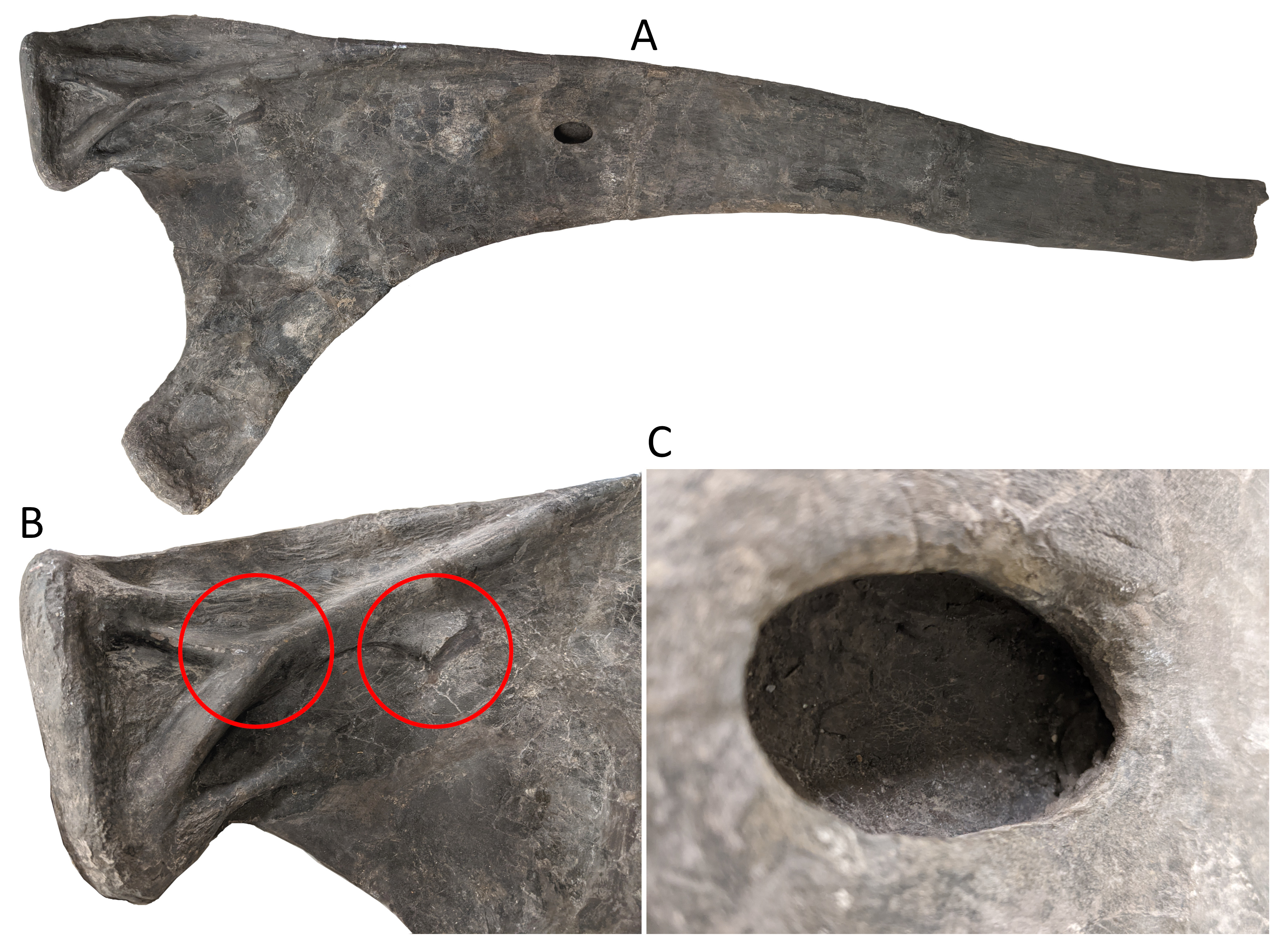
Taylor and Wedel 2023:Figure 2. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, right dorsal rib “Rib A” in posterior view with proximal to the left. A1, the whole proximal half of the rib; a distal portion also exists, of similar length but without features relevant to this study; A2, close-up of the tuberculum, highlighting the complex network of support structures that show signs of speculative reconstruction. Circles highlight two possible sites of the “second tubercle” referred to by Riggs (1901: 549, 1903: 303, 1904: 239) based on Marsh’s illustration (1896: figs. 7, 8), reproduced here in Fig. 4; A3, close-up of the pneumatic foramen in the shaft of the rib, showing natural bone texture around the margin and no indication of breakage. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
This one is visible at the bottom of the photo, proximal end to the bottom, but flipped over from the way it now rests in the collection, so the pneumatic opening is not apparent. There’s an interesting “folded over” ridge running down the anterior(*) face of the proximal part of the shaft.
(*) Assuming we were right in interpreting the available face of the rib as posterior in our paper.
Now, Rib B:

Taylor and Wedel 2023:Figure. 3. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, left dorsal rib “Rib B”. A1, the whole rib, posterior face in proximal view. Foreshortening makes the shaft look shorter and narrower than it actually is: the position of the rib between two shelves makes it impossible to photograph in true posterior view; A2, close-up of the pneumatic opening in the tuberculum in medial view, with anterior to the bottom; A3, red-cyan anaglyph of the same, indicating the form and depth of the fossa. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
This is visible on the left side of the image, close to the vertebral column, with the proximal end to the top. It has the same (posterior, we think) face upwards as is available in the collection, and you can make out the pneumatic opening in the tuberculum that we illustrated.
Reference
Pneumatic dorsal ribs in a selection of ornithodiran taxa. Clades that lack pneumatic ribs have been omitted, including non-dinosaurian dinosauromorphs, ornithischians, all early diverging sauropodomorphs, and numerous sauropods. The only included clade for which dorsal rib pneumaticity might be synapomorphic is Titanosauriformes. Phylogenetic relationships of the sauropods are based on Mannion et al. (2013) for titanosauriforms (note that the position of Brontomerus is uncertain), Tschopp et al. (2015) for diplodocoids, and Zhang et al. (2022) for Xinjiangtitan. Ribs are not shown to scale. Ribs traced from Butler et al. (2009:fig. 1b, Raeticodactylus), Campana (1875:fig. 8, Gallus), Madsen Jr. and Welles (2000:plate 19, Ceratosaurus), Zhang et al. (2022:fig. 14, reversed, Xinjiangtitan), a photo of WDC-DMJ-021-134 provided by David Lovelace (Supersaurus; see Lovelace et al., 2007), Gilmore (1936:plate 29, reversed, Apatosaurus), Riggs (1904:plate 75, Brachiosaurus), Janensch (1950:fig. 108, reversed, Giraffatitan), Wilson and Upchurch (2009:fig. 21, reversed, Euhelopus), Taylor et al. (2011:fig. 7, Brontomerus), and Curry-Rogers (2009:fig. 30, Rapetosaurus). King et al. (2024:fig. 3).
New paper out today with Logan King, Julia McHugh, and Brian Curtice, on pneumatic ribs in Apatosaurus and Brontosaurus (King et al. 2024).
This one had an unusual gestation. In the summer of 2002 2022 I did a road trip to Utah and western Colorado with my friend and frequent collaborator Jessie Atterholt. We did day trips to other collections, but we used Dinosaur Journey in Fruita as home base, and spent most of our time there. That’s where I first met Logan King, who was then recently graduated from Mike Benton’s lab at Bristol. Logan was spending the summer working for Julia McHugh at the Mygatt-Moore Quarry, and Logan and Julia were writing up MWC 9617, a sauropod rib from Mygatt-Moore with interesting pneumatic features.
Now, I had been interested in pneumatic ribs in sauropods for many years, and I’d amassed a war chest of published examples. But I had to admit to myself that the hypothetical pneumatic rib paper I’d been planning was simply never going to be my top priority, and therefore I was never going to actually start it, much less finish it. Logan and I hit it off right away, and I told him I’d be happy to shove my folder of pneumatic rib examples his way, and if he found it useful, I’d be grateful for an acknowledgment. In the actual event, he and Julia asked me to come on as a coauthor, and we were steadily making progress.
That fall I happened to be at Research Casting International at the same time as Brian Curtice — we were both there to see Haplocanthosaurus delfsi while it was down off exhibit from the Cleveland Museum. I’d hung out with Brian a lot back in grad school, but with one thing and another we hadn’t seen each other in many years, and those few days at RCI were a welcome opportunity to rekindle our friendship (and start down the path to coauthorship). Brian also got a look at YPM 1980, the holotype skeleton of Brontosaurus excelsus, while it was at RCI for a remount. Lo and behold, he found unmistakable pneumatic cavities in two of the dorsal ribs of YPM 1980.

A, left rib I, and B, right rib II of YPM 1980, the holotype of Brontosaurus excelsus, in posterior view. King et al. (2024: fig. 2).
That’s pretty awesome for a few reasons. We already knew that the dorsal ribs could be pneumatic in Apatosaurus louisae, because one of the ribs of CM 3018 has a nice round pneumatic cavity. But there was no solid evidence of costal pneumaticity in Brontosaurus. Marsh (1896) figured a rib with pneumatic cavities and claimed it for Brontosaurus, but without a specimen number the referral was uncertain. Turns out there is costal pneumaticity in Brontosaurus, and not just any bronto, but the ur-brontosaur itself, YPM 1980. And in 143 years, no-one had clocked it (there’s a lot of that going around). It seemed silly to write up a pneumatic rib of Apatosaurus from Mygatt-Moore and not mention the newly-discovered rib pneumaticity in YPM 1980, so we brought Brian in on the project. The manuscript went through a genuinely constructive review process at JVP, and we were revising the text and figs last fall.
While I had the apatosaur rib pneumaticity paper with Logan, Julia, and Brian going on one burner, Mike went to Chicago, decided that Brachiosaurus ribs were worth looking at after all (full story here), and went and wrote an entire paper on them in essentially no time. So after deciding in July of 2022 that I was never going to get around to my sauropod rib paper and I should hand it off to someone else (which was absolutely the right decision), a mere 14 months later I found myself working on two sauropod rib papers simultaneously. But they were on different taxa and had somewhat different focuses, so I made my junior author contributions to both and tried not to let Brachiosaurus step on Apatosaurus’s toes. (In particular, Mike and I didn’t talk much about pneumatic ribs outside of Brachiosauridae because there was already a broader survey in Logan’s manuscript.) Brach flew through review and into print just before year’s end (Taylor and Wedel 2023), and now the apatosaurines have lumbered over the finish line. I’m proud of both papers, and very happy to have them out in the world.

Proximal rib head that compromises MWC 9617 in posterior view. The inset image depicts a line drawing of the section of the rib that preserves pneumatic fossae within the rib canal sulcus. Abbreviations: cp, capitulum; I, proximal pneumatic fossa; II, middle pneumatic fossa; III, distal pneumatic fossa; t, tuberculum. Scale bar equals 5 cm. King et al. (2024:fig. 1).
MWC 9617 is an interesting specimen, with a series of same-sized fossae running down the postero-medial side, inside a long sulcus. That’s the side of the rib where the intercostal nerve, artery, and vein would have run — because that’s where they run in all tetrapods — but that neurovascular bundle doesn’t usually sit in a sulcus in sauropod ribs (the same neurovascular bundle does sit in a groove on the underside of human ribs). Those fossae are too smooth and too regular to be pathological. Pneumatic excavations that far down the rib shaft are unusual but not unprecedented — some of the ribs of Paluxysaurus and the Wyoming Supersaurus have pneumaticity about that far distally, and then there’s the weird lonely foramen in the one rib of Brachiosaurus that Riggs (1904) did illustrate. And sometimes pneumatic diverticula do create repeated excavations that look almost identical; one of my favorite examples is the series of pneumatic foramina on the right side of the centrum in a cervical vertebra of (perhaps fittingly) Paluxysaurus. So this certainly looks like a large pneumatic excavation, which we might call a fossa or a sulcus, containing smaller subfossae excavated at regular intervals. That’s pretty cool, because although that general mode of pneumatization turns up now and then in vertebrae, nobody’s documented it in a rib before.

C5? of Paluxysaurus in right lateral view, traced from a photo I took at the Fort Worth Museum of Science and History back in 1990s. I should do a separate post just on this vert sometime — the pneumatic excavations on the left side of the centrum are completely different.
We think that MWC 9617 is a rib of Apatosaurus louisae, for a couple of reasons. One, A. louisae is the most common sauropod at Mygatt-Moore by a wide margin, so any given rib from MMQ is more likely to belong to Apatosaurus than to anything else. The other sauropods known from MMQ so far are Camarasaurus and an indeterminate diplodocine (Foster et al. 2018) — and no pneumatic ribs have ever been described for either Camarasaurus or any of the Morrison diplodocines. (That in itself is pretty weird, given that Diplodocus and especially Barosaurus have pretty complex and extensive vertebral pneumaticity. How did a thicc boi like Apatosaurus beat them to the punch on pneumatizing ribs?) Anyway, it’s more parsimonious that the pneumatic rib from the apatosaur-dominated quarry belongs to Apatosaurus, for which pneumatic ribs are already known, than that it belongs to Camarasaurus or a diplodocine, for which it would be a world first. Bottom line, if we’re wrong, that’s even more exciting.
What’s next? At some point, more stuff from Mygatt-Moore! Jessie and I made Dinosaur Journey home base for our 2022 research trip because neither of us had ever gotten more than one day at a time in that collection. With a whole week to play there, and Julia and Logan to show us weird stuff, we made a LOT of progress, and found some stuff even I didn’t expect. Watch this space.
If you’re around sauropod material, look at ribs. Even the ones that were described in the 1800s may surprise you. Describing pneumaticity is everyone’s business — if you see something, say something!
References
- Foster, J., Hunt-Foster, R., Gorman, M., Trujillo, K., Suarez, C., McHugh, J., Peterson, J., Warnock, J. and Schoenstein, H., 2018. Paleontology, taphonomy, and sedimentology of the Mygatt-Moore Quarry, a large dinosaur bonebed in the Morrison formation, western Colorado—implications for Upper Jurassic dinosaur preservation modes. Geology of the Intermountain West 5:23-93.
- King, J.L., McHugh, J.B., Wedel, M.J., and Curtice, B. 2024. A previously unreported form of dorsal rib pneumaticity in Apatosaurus (Dinosauria: Sauropoda) and implications for pneumatic variation among diplodocid dorsal ribs. Journal of Vertebrate Paleontology. DOI: 10.1080/02724634.2024.2316665
- Marsh, O.C. 1896. The Dinosaurs of North America. 16th annual report of the U. S. Geological Survey, 1894-95, pt. I. US Government Printing Office, Washington, D.C.
- Riggs, Elmer S. 1904. Structure and relationships of opisthocoelian dinosaurs. Part II, the Brachiosauridae. Field Columbian Museum, Geological Series 2(6):229-247, plus plates LXXI-LXXV.
- Taylor, Michael P., and Mathew J. Wedel. 2023. Novel pneumatic features in the ribs of the sauropod dinosaur Brachiosaurus altithorax. Acta Palaeontologica Polonica 68(4): 709–718. doi:10.4202/app.01105.2023
Opportunism, helpfulness, disappointment, persistence: the genesis of the Brachiosaurus rib paper
January 12, 2024
Just a quick post about the genesis of the Brachiosaurus rib paper (Taylor and Wedel 2023) that I wrote about at the very end of last year. Although this is in some respects a minor paper, I’m fond of it because it fell into place so quickly and easily.
Taylor and Wedel 2023:Figure. 3. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, left dorsal rib “Rib B”. A1, the whole rib, posterior face in proximal view. Foreshortening makes the shaft look shorter and narrower than it actually is: the position of the rib between two shelves makes it impossible to photograph in true posterior view; A2, close-up of the pneumatic opening in the tuberculum in medial view, with anterior to the bottom; A3, red-cyan anaglyph of the same, indicating the form and depth of the fossa. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
The background: from 14–22 August last year I was in Chicago with my day-job. The work events finished on the 21st and it happened that the best-value flight back home from O’Hare wasn’t until nearly 7pm on the 22nd. So I had the best part of a day free in Chicago.
When I’d been in California with Matt on the previous week, we’d seen something in the LACM public gallery[1] that I wanted to follow up. I only realised quite late in the day that my late flight gave me an opportunity to look at material in Chicago’s Field Museum of Natural History. So it wasn’t until 19 August that I dropped a line to Bill Simpson, the Field Museum’s fossil-reptile collection manager, asking if I could visit in three days. (Actually it’s even worse than that: I got my dates wrong and asked if I could visit in four days, then had to go back and change it!)
Well, Bill was super-helpful, just as he had been when Matt and I visited way back in 2005. He arranged to let me into the collection at 8:30am and then leave me free to look through the material, armed with a spreadsheet of what’s where. I really can’t emphasize enough how consistently helpful I’ve found pretty much all museum staff when I’ve visited anywhere — contra the unfair stereotype of curators being obstructive or difficult — but the Field Museum has gone above and beyond for me on both my visits.
Unfortunately, the mystery purpose of my visit[1] was a bust: I was disappointed to find that the thing I was looking for wasn’t there. But the day wasn’t a bust. When I’d told Matt I’d been able to arrange the visit, he’d written to me:
I know you’ll be moving quickly but I have a request […] AFAIK the only rib of BOBA that’s ever been figured by anyone, and the only photo of that rib, is the one included by Riggs that shows the proximal 1/4 or so with a pneumatic foramen on the shaft. Any photos of that rib or any others would be super helpful, especially if they show pneumatic features. […] If you could get photos without scale bars in the way, and if possible measurements of foramen dimensions, that would be fantastic, and probably worth a short note somewhere, since the foramen on the shaft of Riggs’s rib is the only costal pneumaticity ever documented in Brachiosaurus.
(The photo that Matt attached to that email was one that we took in 2005 of what we’re now calling “Rib B” — not Rib A, the one that Riggs had illustrated. Here it is.)

Brachiosaurus altithorax holotype FMNH P 25107, proximal portion of ?left rib “Rib B” in ?posterior view, with dorsal to the top right. Photo taken in 2005, when we were young and stupid.
I wrote back to Matt:
We hardly need to write a short note on BOBA’s costal pneumaticity since we already mention it in [an in-progress manuscript]. We just need to substitute in a better image and some measurements.
Idiotically, I needed Matt to persuade me that this was something worth looking into. It’s worth quoting his message at length, since it contains all the seeds of what eventually became this paper:
Sorry, didn’t explain myself well re: BOBA costal pneumaticity. In Giraffatitan, the only (documented) pneumatic features in the dorsal ribs are foramina that open into the tuberculum — on both sides! See attached cap from Janensch [which turns up in our new paper’s Figure 5B]. And in fact that is one of the most common places for costal pneumaticity to manifest in sauropods, the other being on the web of bone that connects the tuberculum to the capitulum (as in Brontomerus [reproduced in the new paper’s Figure 5E]).
BOBA has been an anomaly in only having pneumaticity documented a decent distance down the shaft; I’ve found other sauropods that have lines of fossae or foramina that extend down about that far (Supersaurus and Paluxysaurus if you’re curious), but no others that have a long, bare shaft and then BOOM a totally isolated pneumatic foramen. I don’t doubt that foramen is real, but right now it’s the only form of costal pneumaticity in BOBA that anyone other than you and me knows about (or, if others have noticed the proximal pneumatic features, they haven’t said anything in public). Documenting more proximal pneumatic features will be valuable, both for showing that BOBA does have those features in common with other brachiosaurids and other sauropods more generally, and for showing that a single individual can have a diversity of pneumatic features at different distances down the rib. Not terribly surprising for us, but probably more so for people who are less well-acquainted with rampaging diverticula on the loose.
Well, obviously Matt was right, and I spent the bulk of my Field Museum visit with those ribs. The new paper in Acta Pal. Pol. is the result. It’s Taylor and Wedel rather than Wedel and Taylor because I was the one who visited the fossils, photographed and measured them, wrote the text and prepared the illustrations. But in a deeper sense, it’s Matt’s paper — it would never have happened at all without his insight and insistence.
Once I was convinced, it didn’t take long to get the paper written up and illustrated: it was submitted on 1 September, eight days after I got home from America. It had an unusually straightforward path through peer-review, thanks to positive and constructive reviews from Jerry Harris, Virginia Zurriaguz and Pat O’Connor. And it went pretty quickly through production, as the people at Acta Pal. Pol. wanted the issue containing it to come out in 2023. The result is that it took four months and a week from seeing the specimen to publication of the paper.
So that’s how we ended up with this new paper: my opportunism in sneaking a collections visit while I had a few spare hours in Chicago; Bill Simpson’s helpfulness in making that happen at very short notice; my disappointment in not finding what I’d been looking for in the collections[1]; and Matt’s persistence in pushing me to properly look at the pneumaticty in the BOBA ribs.
References
Notes
[1] I’m keeping the powder dry on this for now, but we’ll probably talk about it soon.
The ribs of Brachiosaurus: stranger than you thought
December 30, 2023
As we’ve often observed, it’s a funny thing that incredibly well-known dinosaur specimens can sit around for decades, or for more than a century, before someone notices something fascinating about them. One lesson to learn from this is the importance of collections — their creation, maintenance and accessibility. Another is of course to always look at the fossils we see.
In another iteration of this old theme, today sees the publication of Taylor and Wedel (2023), a short paper on pneumatic features in the dosal ribs of our old friend FMNH PR 25107, the holotype of Brachiosaurus altithorax.
Taylor and Wedel 2023:Figure 2. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, right dorsal rib “Rib A” in posterior view with proximal to the left. A1, the whole proximal half of the rib; a distal portion also exists, of similar length but without features relevant to this study; A2, close-up of the tuberculum, highlighting the complex network of support structures that show signs of speculative reconstruction. Circles highlight two possible sites of the “second tubercle” referred to by Riggs (1901: 549, 1903: 303, 1904: 239) based on Marsh’s illustration (1896: figs. 7, 8), reproduced here in Fig. 4; A3, close-up of the pneumatic foramen in the shaft of the rib, showing natural bone texture around the margin and no indication of breakage. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
Here’s the thing about this rib, which Riggs illustrated in two of his papers — the initial brief description of Brachiosaurus (Riggs 1903:figure 6) and the subsequent monographic osteology (Riggs 1904:plate LXXV:figure 5). It has a pneumatic condition that has not been documented in any other sauropod specimen.
The pneumatic opening is part way down the shaft — about 60 centimeters down from the tuberculum. But there are no other pneumatic features more proximally on the same face of the rib. (We don’t know what’s on the other face: it’s sitting in a plaster half-jacket, and flipping it would not be trivial.)
What does this mean? The seven-location schema of Wedel and Taylor (in revision) predicts that pneumatic features in costal elements would follow vascular foramina from the segmental and intercostal arteries. The segmental arteries pass behind the ribs on their circuit of the centrum, vacularizing the posterior aspect of the proximal portion of the ribs, the tubercula and capitula and region between them — and so providing channels for pneumatization in these regions. Meanwhile, intercostal arteries extend along and beyond the length of the rib shaft, providing opportunities for vascularization and subsequent pneumatization.
But while it’s pretty common to see pneumatization of the proximal portions of ribs, pneumatization of the shafts — likely by diverticula following the intercostal arteries — is hardly ever seen. In fact “Rib A” of the Brachiosaurus altithorax holotype provides the only documented occurrence, and that has only been recognized 120 years after the initial description.
All of this is more evidence for the opportunistic and random behaviour of pneumatizing diverticula. They always have the possibility of passing along the length of rib shafts and pneumatizin them — but either they rarely extend along the intercostal arteries, or if they do then they rarely excavate the bone that they are adjacent to. Why? We have no idea. It seems to be just the way the dice fall.
References
- Riggs, Elmer S. 1903. Brachiosaurus altithorax, the largest known dinosaur. American Journal of Science 15(4):299-306.
- Riggs, Elmer S. 1904. Structure and relationships of opisthocoelian dinosaurs. Part II, the Brachiosauridae. Field Columbian Museum, Geological Series 2(6):229-247, plus plates LXXI-LXXV.
- Taylor, Michael P., and Matthew J. Wedel. 2023. Novel pneumatic features in the ribs of the sauropod dinosaur Brachiosaurus altithorax. Acta Palaeontologica Polonica 68(4): 709–718. doi:10.4202/app.01105.2023
In mammals — certainly the most-studied vertebrates — regional differentiation of the vertebral column is distinct and easy to spot. But things aren’t so simple with sauropods. We all know that the neck of any tetrapod is made up of cervical vertebrae, and that the trunk is made up of dorsal vertebrae (subdivided into thoracic and lumbar vertebrae in the case of mammals). But how do we tell whether a given verebra is a posterior cervical or an anterior dorsal?
Here two vertabrae: a dorsal vertebra (D3) and a cervical vertebra (C13) from CM 84, the holotype of Diplodocus carnegii, modified from Hatcher (1901: plates III and VII):

It’s easy to tell these apart, even when as here we have only lateral-view images: the dorsal vertebra is tall, its centrum is short, its neural spine is anteroposteriorly compressed and its parapophysis is up on the dorsal half of the centrum; but the cervical vertebra is relatively low, its centrum is elongated, its neural spine is roughly triangular and its parapophysis hangs down well below the centrum (and has a cervical rib fused to it and the diapophysis).
But things get trickier in the shoulder region because, in sauropods at least, the transition through the last few cervicals to the first few dorsals is gradual — the vertebrae become shorter, taller and broader — and tends to have no very obvious break point. In this respect, they differ from mammals, in which the regional differentiation of the spinal column is more abrupt. (Although even here, things may not be as simple as generally assumed: for example, Gunji and Endo (2016) argued that the 1st thoracic vertebra of the giraffe behaves functionally like an 8th cervical.)
So here are those two vertebrae in context: the sequence D3 D2 D1 C15 C14 C13 in CM 84, the holotype of Diplodocus carnegii, modified from Hatcher (1901: plates III and VII):

Given that the leftmost is obviously a dorsal and the rightmost obviously a cervical, where would you place the break-point?
The most usual definition seems to be that the first dorsal vertebra is the first one that has a free rib, i.e. one not fused to the vertebra: in the illustration above, you can see that the three cervicals on the right all have their cervical ribs fused to their diapophyses and parapophyses, and the three dorsals on the left do not. This definition of the cervical/dorsal distinction seems to be widely assumed, but it is rarely explicitly asserted. (Does anyone know of a paper that lays it out for sauropods, or for dinosaurs more generally?)
But wait!
Hatcher (1903:8) — the same dude — in his Haplocanthosaurus monograph, writes:
The First Dorsal (Plate I., Fig. 1). […] That the vertebra now under consideration was a dorsal is conclusively shown not by the presence of tubercular and capitular rib facets showing that it supported on either side a free rib, for there are in our collections of sauropods, skeletons of other dinosaurs fully adult but, with the posterior cervical, bearing free cervical ribs articulating by both tubercular and capitular facets as do the ribs of the dorsal region. The character in this vertebra distinguishing it as a dorsal is the broadly expanded external border of the anterior branch of the horizontal lamina [i.e. what we would now call the centroprezygapophyseal lamina]. This element has been this modified in this and the succeeding dorsal, no doubt, as is known to be the case in Diplodocus to give greater surface for the attachment of the powerful muscles necessary for the support of the scapula.
Hatcher’s illustrations show this feature, though they don’t make it particularly obvious: here are the last two cervicals and the first dorsal, modified from Hatcher (1903:plate I), with the facet in question highlighted in pink: right lateral view at the top, then anterior, and finally posterior view at the bottom. (The facet is only visible in lateral and anterior views):

Taken at face value, Hatcher’s words here seem to imply that he considers the torso to begin where the scapula first lies alongside the vertebral column. Yet if you go back to the Diplodocus transition earlier in this post, a similar scapular facet is not apparent in the vertebra that he designated D1, and seems to be present only in D2.
Is this scapular-orientation based definition a widespread usage? Can anyone point me to other papers that use it?
Wilson (2002:226) mentions a genetic definition of the cervical/dorsal distinction
Vertebral segment identity may be controlled by a single Hox gene. The cervicodorsal transition in many tetrapods, for instance, appears to be defined by the expression boundary of the Hoxc-6 gene.
But this of course is no use in the case of extinct animals such as sauropods.
So what’s going on here? In 1964, United States Supreme Court Justice Potter Stewart, in describing his threshold test for obscenity, famously said “I shall not today attempt further to define the kinds of material I understand to be embraced within that shorthand description, and perhaps I could never succeed in intelligibly doing so. But I know it when I see it.” Is that all we have for the definition of what makes a vertebra cervicals as opposed to dorsal? We know it when we see it?
Help me out, folks! What should the test for cervical-vs-dorsal be?
References
- Gunji, Mego, and Hideki Endo. 2016. Functional cervicothoracic boundary modified by anatomical shifts in the neck of giraffes. Royal Society Open Science 3:150604. doi:10.1098/rsos.150604
- Hatcher, Jonathan B. 1901. Diplodocus (Marsh): its osteology, taxonomy and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1:1-63 and plates I-XIII.
- Hatcher, J. B. 1903b. Osteology of Haplocanthosaurus with description of a new species, and remarks on the probable habits of the Sauropoda and the age and origin of the Atlantosaurus beds; additional remarks on Diplodocus. Memoirs of the Carnegie Museum 2:1-75 and plates I-VI.
- Wilson, Jeffrey A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136:217-276.
How fat is an elephant?
May 29, 2013
We jumped the gun a bit in asking How fat was Camarasaurus? a couple of years ago, or indeed How fat was Brontosaurus? last year. As always, we should have started with extant taxa, to get a sense of how to relate bones to live animals — as we did with neck posture.
So here we go. I give you a herd of Indian elephants, Elephas maximus (from here):

You will notice, from this conveniently-close-to-anterior view, that their torsos bulge out sideways, much further than the limbs.
Now let’s take a look at the skeleton of the same animal in the Oxford University Museum of Natural History (downloaded from here but for some reason the photo has now gone away):

The rib-cage is tiny. It doesn’t even extend as far laterally as the position of the limb bones.
(And lest you think this is an oddity, do go and look at any mounted elephant skeleton of your choice, Indian or African. They’re all like this.)
What’s going on here?
Is Oxford’s elephant skeleton mounted incorrectly? More to the point, are all museums mounting their elephants incorrectly? Do elephants’ ribs project much more laterally in life?
Do elephants have a lot of body mass superficial to the rib-cage? If so, what is that mass? It’s hard to imagine they need a huge amount of muscle mass there, and it can’t be guts. Photos like this one, from the RVC’s televised elephant dissection on Inside Nature’s Giants, suggest the ribs are very close to the body surface:

I’m really not sure how to account for the discrepancy.
Were sauropods similarly much fatter than their mounted skeletons suggest? Either because we’re mounting their skeletons wrongly with the ribs too vertical, or because they had a lot of superficial body mass?
Consider this mounted Camarasaurus skeleton in the Dinosaur Hall at the Arizona Museum of Natural History (photo by N. Neenan Photography, CC-BY-SA):

Compare the breadth of its ribcage with that of the elephant above, and then think about how much body bulk should be added.
This should encourage palaeoartists involved in the All Yesterdays movement to dramatically bulk up at least some of their sauropod restorations.
It should also make us think twice about our mass estimates.
I’ve recently written about my increasing disillusionment with the traditional pre-publication peer-review process [post 1, post 2, post 3]. By coincidence, it was in between writing the second and third in that series of posts that I had another negative peer-review experience — this time from the other side of the fence — which has left me even more ambivalent about the way we do things.
On 17 July I was asked to review a paper for Biology Letters. Having established that it was to be published as open access, I agreed, was sent the manuscript, and two days later sent a response that recommended acceptance after only minor revision. Eleven days later, I was sent a copy of the editor’s decision — a message that included all three reviewers’ comments. I can summarise those reviewers’ comments by directly quoting as follows:
Revewer 1: “It is good to have this data published with good histological images. I have only minor comments – I think the ms should generally be accepted as it is.”
Reviewer 2 (that’s me): “This is a strong paper that brings an important new insight into a long-running palaeobiological issue […] and should be published in essentially its current form.”
Reviewer 3: “This manuscript reports exciting results regarding sauropod biomechanics […] The only significant addition I feel necessary is to the concluding paragraph.”
So imagine my surprise when the decision letter said:
I am writing to inform you that your manuscript […] has been rejected for publication in Biology Letters.
This action has been taken on the advice of referees, who have recommended that substantial revisions are necessary. With this in mind we would like to invite a resubmission, provided the comments of the referees are taken into account. This is not a provisional acceptance.
The resubmission will be treated as a new manuscript.
I can’t begin to imagine how they turned three “accept with very minor revisions” reviews into “your manuscript has been rejected … on the advice of referees, who have recommended that substantial revisions are necessary”.
In fact, let’s dump the “I can’t imagine how” euphemism and say it how it is: “reviewers recommended substantial revisions” is an outright lie. The reviewers recommended no such thing. The rejection can only be because it’s what the editor wanted to do in spite of the reviewers’ comments not because of them. It left me wondering why I bothered to waste my time offering them an opinion that they were only ever going to ignore.
—
Then six days ago I heard from the lead author, who had just had a revised version of the same manuscript accepted. (It had not come back to me for review, as the editor had said would happen with any resubmission).
The author wrote to me:
The paper will be published (open access) at the 3rd of Octobre. When I had submitted the corrected version of the ms acceptance was only a formality. So [name] was right, they just want to keep time between submission and publishing date short.
Well. We have a word for this. We call it “lying”. When the editor wrote “your manuscript […] has been rejected for publication in Biology Letters … With this in mind we would like to invite a resubmission … This is not a provisional acceptance. The resubmission will be treated as a new manuscript”, what she really meant was “your manuscript […] has been provisionally accepted, please sent a revision. The resubmission will not be treated as a new manuscript”.
I find this lack of honesty disturbing.
Because we’re not talking here about some shady, obscure little third-world publisher that no-one’s ever heard of with fictional people on the editorial board. We’re talking about the Royal Freaking Society of London. We’re talking about a journal (Biology Letters) that was calved off a journal (Proceedings B) that emerged from the oldest continuously published academic journal in the world (Philosophical Transactions). We’re talking about nearly three and a half centuries of academic heritage.
And they’re lying to us about their publication process.
When did they get the idea that this was acceptable?
And what else are they lying to us about? Can we trust (for example) that when editors or members submit papers, they are subjected to the same degree of rigorous filtering as every other submission? I would have assumed that, yes, of course they do. But I just don’t know any more.

Sampled specimens, sampling locations and cross sections of sauropod cervical ribs. (a) Anterior neck of Brachiosaurus brancai (Museum für Naturkunde, Berlin) with hyperelongated and overlapping cervical ribs. (b) Three cross sections were taken along the proximal part of the posterior process of a left mid-neck cervical rib of Mamenchisaurus sp. (SIPB 597) in ventral view. Note the medially pointed ventral part of the cervical rib. (c) Seven cross sections were taken along the left ninth cervical rib of B. brancai (MB.R.2181.90), which is figured in lateral view. (d) Neck of Diplodocus carnegi (cast in the Museum für Naturkunde, Berlin) with short cervical ribs. (e) Six cross sections were taken along the right mid-neck cervical rib of cf. Diplodocus sp. (Sauriermuseum Aathal, Aathal HQ2), which is figured in ventral view. Note the morphological differences of this cervical rib when compared with the hyperelongated cervical rib of B. brancai. (Klein et al. 2012:figure 1)
The paper in question is Klein et al.’s (2012) histological study confirming that the bony cervical ribs of sauropods are, as we suspected, ossified tendons — as we assumed in our recently arXiv’d sauropod-neck paper. I am delighted to be able to say that it is freely available. At the bottom of the first page, it says “Received 21 August 2012; Accepted 13 September 2012”, for a submission-to-acceptance time of 23 days. But I know that the initial submission — and remember, the final published version is essentially identical to that initial submission — was made before 17 July, because that’s when I was asked to provide a peer-review. Honest reporting would give a submission-to-acceptance time of 58 days, which is two and a half times as long as the claimed figure.
Now the only reason for a journal to report dates of submission and acceptance at all is to convey the speed of turnaround, and lying about that turnaround time completely removes any utility those numbers might have. It would be better to not report them at all than to fudge the data.
This is another way that the high-impact fast-turnaround publishing system is so ridiculously gamed that it actually hurts science. We have the journal lying to authors about the status of their manuscripts so that it can then lie to the readers about its turnaround times. That’s deeply screwed up. And it’s hard for authors to blow the whistle — they don’t want to alienate the journals and the editors who have some veto power over their tenure beans, and reviewers don’t usually have all the information. The obvious solution is to make the peer-review process more open, and to make editorial decisions more transparent.
That, really, is only what we’d respect from the Royal Society. Isn’t it?
—
Note. Nicole Klein did not know I was going to post about this. I want to make that clear so that no-one at the Royal Society thinks that she or any of her co-authors is making trouble. All the trouble is of my making (and, more to the point, the Royal Society’s). Someone really has to shine a light on this misbehaviour.
Update (12 March 2014)
I should have noted this before, but on 10 May 2013, the Royal Society sent me an update, explaining some improvements in their process. But as noted in my write-up, it doesn’t actually solve the problem. Doing so would simply require giving three dates: Received, Revised and Accepted. But as I write this, new Proc. B articles still only show Received and Accepted dates.
Reference
Subsequent posts discuss how this issue is developing:
- We will no longer provide peer reviews for Royal Society journals until they adopt honest editorial policies
- Biology Letters does trumpet its submission-to-acceptance time
- Lying about submission times at other journals?
- Discussing Biology Letters with the Royal Society
- The Royal Society has taken some steps to improving reporting of submit/resubmit/accept times
- Fumbling towards transparency: the Royal Society’s “reject & resubmit” and submitted/published dates


{kind=link}